

Text als pdf
Pelecaniformes
Die Pelecaniformes sind die dritte Ordnung innerhalb der Pelicanimorphae. Die beiden anderen Ordnungen, die Ciconiiformes und Suliformes lernten wir in der letzten Episode kennen. Zu den Pelecaniformes gehören die Pelikane (Pelecanidae), der Schuhschnabel (Balaeniceps rex), der Hammerkopf (Scopus umbretta), die Reiher (Ardeidae) sowie die Ibisse und Löffler (Threskiornithidae). Bis vor wenigen Jahren hatte die Ordnung eine fast völlig andere Zusammensetzung und neben den Pelikanen wurden die Tölpel (Sulidae), die Fregattvögel (Fregatidae), die Tropikvögel (Phaethontidae), die Kormorane (Phalacrocoracidae) und die Schlangenhalsvögel (Anhingidae) in die Pelecaniformes gestellt. In dieser Zusammensetzung wurden die Pelecaniformes im Deutschen Ruderfüßer genannt, nach dem Bau der Füße, bei denen alle vier Zehen durch eine Schwimmhaut verbunden sind. Die Ähnlichkeiten zwischen Pelikanen und den anderen Familien beruhen jedoch lediglich auf Konvergenz, molekularbiologische Befunde wie DNA-Vergleiche von mitochondrialer und nukleärer (im Zellkern befindlicher) DNA sprechen deutlich gegen eine Verwandtschaft (Hackett et al. 2008, Jarvis et al. 2014, Kuhl et al. 2021, Prum et al. 2015, Kuramoto et al. 2015, Sangster & Mayr 2021, Sangster et al. 2022, Suh et al. 2005, Wu et al. 2024, Winkler et al. 2015, Kimball et al. 2013, Braun & Kimball 2021, Abb. 1).
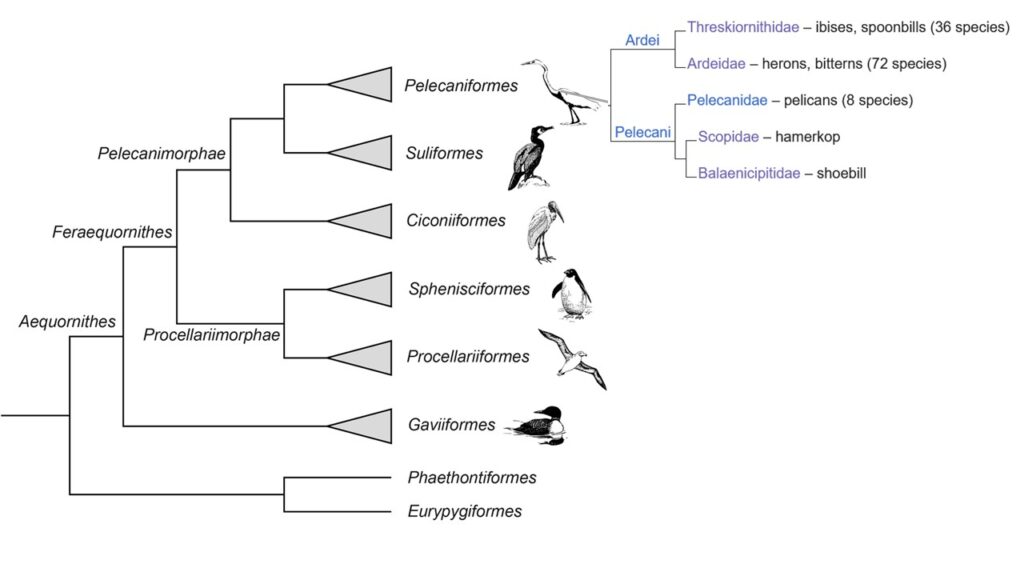
Abb. 1: Kladogramm der Aequornithes
Die nächsten Verwandten der Pelikane sind der Schuhschnabel (Balaeniceps rex) und der Hammerkopf (Scopus umbretta), zwei Vögel, die im tropischen Afrika heimisch sind und in der traditionellen Systematik zu den Schreitvögeln (Ciconiiformes) gerechnet wurden. Schwestergruppe der von allen drei Taxa gebildeten Klade sind die Reiher (Ardeidae) und die Ibisse und Löffler (Threskiornithidae). Schwestergruppe der von allen fünf Taxa gebildeten Klade sind die übrig gebliebenen Ruderfüßer. Um wieder zu monophyletischen Taxa zu kommen, ordnet das International Ornithological Committee alle Familien der Schreitvögel bis auf die Störche (Ciconiidae) den Pelecaniformes zu. Die Tropikvögel bilden jetzt eine eigenständige Ordnung, die Phaethontiformes, die nicht zu den Aequorntihes, sondern zusammen mit Kagu und Sonnenralle zu den Eurypigimorphae gezählt werden, und die übrigen Ruderfüßer werden in die Ordnung Suliformes gestellt.
Ibisse und Löffler (Threskiornithidae)
Ibisse und Löffler umfassen zwei Vogeltypen, die ursprünglich als eigene Unterfamilien angesehen wurden. Sie sind die basalste Klade der Pelecanifomres. Insgesamt gibt es 36 Arten in 13 Gattungen. Lange Zeit wurden sie zusammen mit den Störchen zu den Schreitvögeln (Ciconiiformes) gestellt. Insbesondere die Nimmersatte weisen viele Ähnlichkeiten mit den Ibissen auf. Mittlerweile weiß man, dass diese Ähnlichkeiten auf konvergente Evolution beruhen. Ihre Schwestergruppe sind die Reiher (van Tuinen et al. 2001, Hackett et al. 2008, Jarvis et al. 2014, Kuhl et al. 2021, Prum et al. 2015, Kuramoto et al. 2015, Sangster & Mayr 2021, Sangster et al. 2022, Suh et al. 2005, Wu et al. 2024, Winkler et al. 2015, Kimball et al. 2013, Braun & Kimball 2021, Abb. 2).
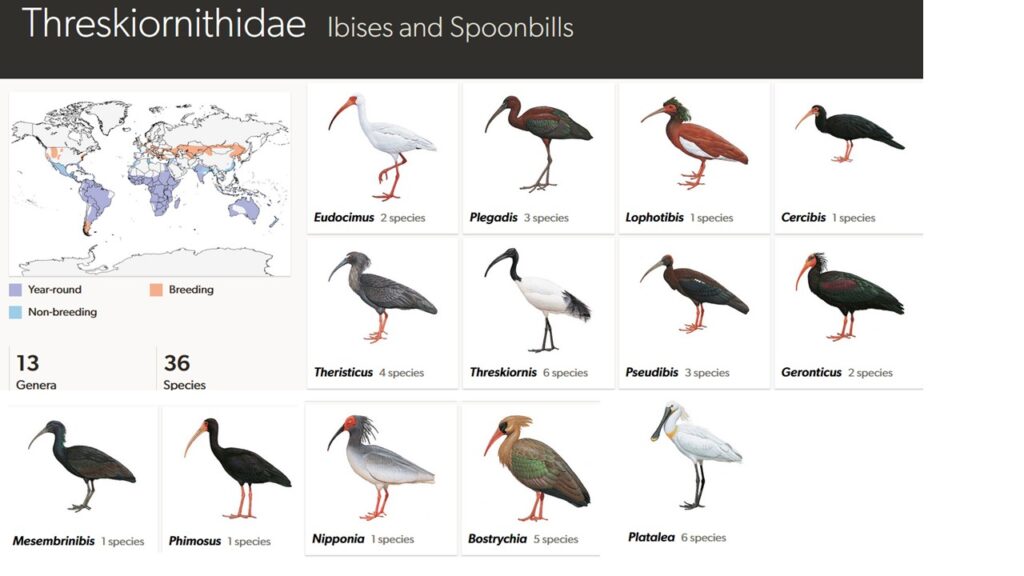
Abb. 2: Ibisse
Ibisse und Löffler sind 50 bis 110 cm lang, haben einen gedrungenen Körper sowie einen langen Hals und lange Beine. Die Beine enden in einem anisodaktylen Fuß mit langen Zehen; Schwimmhäute sind nur basal vorhanden und kaum auszumachen. Die Flügel sind lang und breit und ermöglichen einen kräftigen, schnellen Flug. Charakteristisch für Ibisse ist der lange, abwärts gebogene Schnabel. Löffler dagegen haben einen langen, an der Schnabelspitze auffällig verbreiterten und abgeplatteten Schnabel. Es gibt weiß, braun, schwarz und rot gefiederte Arten. Braune und schwarze Farben kommen allerdings nur bei den Ibissen vor. Vom Rosalöffler abgesehen, weisen die Löffler ein weißes Gefieder auf. Zudem haben alle Arten unbefiederte Hautpartien, meistens im Gesichtsbereich. Einen Geschlechtsdimorphismus gibt es – abgesehen von einem geringfügigen Größenunterschied – nicht. Allerdings gibt es einen jahreszeitlichen Wechsel: zur Brutzeit erscheinen Gefieder und nackte Hauptpartien leuchtender.
Ibisse und Löffler kommen fast weltweit vor, mit einem Verbreitungsschwerpunkt in den Tropen. Der typische Lebensraum sind Seen oder langsam fließende Flüsse, sowohl in offenen Landschaften als auch in dichten Regenwäldern. Einige Arten leben aber auch in Steppen, Savannen und Wüsten. Die wasserbewohnenden Arten fressen Wasserinsekten, Insektenlarven, Kleinkrebse und Mollusken, seltener auch kleine Fische und Amphibien. Die wenigen Arten, die abseits vom Wasser in trockeneren Gegenden leben, ernähren sich hingegen von Heuschrecken, Käfern, Spinnen und Schnecken, seltener auch von Eidechsen, Schlangen und Mäusen (Winkler et al. 2015, 2020a, Del Hoyo 2020, Del Hoyo et al. 1992).
Die meisten Ibisse und Löffler brüten in Kolonien. Zur Balz und in geringerem Maße auch später zur Begrüßung am Nest gibt es eine große Zahl ritualisierter Gesten, zum Beispiel das Senken des Kopfes, das Aneinanderreiben der Schnäbel und Hälse sowie den Austausch von Nistmaterial.
Beide Partner beteiligen sich am Nestbau. Für gewöhnlich wird das Nest in einem Baum gebaut und besteht aus Zweigen, Gräsern und sonstigem Pflanzenmaterial. Dann legt das Weibchen zwei bis fünf, selten bis sieben Eier. Beide Partner brüten für einen Zeitraum von 20 bis 31 Tagen. Wenn die Jungvögel schlüpfen, haben sie einen kurzen, geraden Schnabel. Dieses gemeinsame Merkmal wurde auch als Beweis für die enge Verwandtschaft von Ibissen und Löfflern herangezogen. Schon nach einigen Tagen beginnt der Schnabel mit seinem schnellen Wachstum: Bei den Ibissen wird er lang und gebogen, während sich bei den Löfflern die Verbreiterung herausbildet. Schon im Alter von 16 Tagen hat ein Junglöffler einen Schnabel, der dem der Eltern gleicht.
Löffler wurden lange Zeit als eigene Unterfamilie angesehen und den Ibissen gegenübergestellt. Tatsächlich sind die Ibisse aber paraphyletisch, d. h. einige Ibisse sind mit den Löfflern enger verwandt als mit den restlichen Ibissen, weswegen die Einteilung in diese zwei Unterfamilien aufgegeben wurde. Stattdessen kann man innerhalb der Familie zwei Kladen unterscheiden, eine nur in der Neuen Welt vorkommende, bestehend aus den Gattungen Eudocimus, Phimosus und Theristicus, und eine fast weltweit verbreitete mit den übrigen Gattungen einschließlich der Löffler. Beide Evolutionslinien trennten sich schon vor 39 bis 42 Millionen Jahren voneinander (Chesser et al. 2010, Ramirez et al. 2013, Abb. 3).
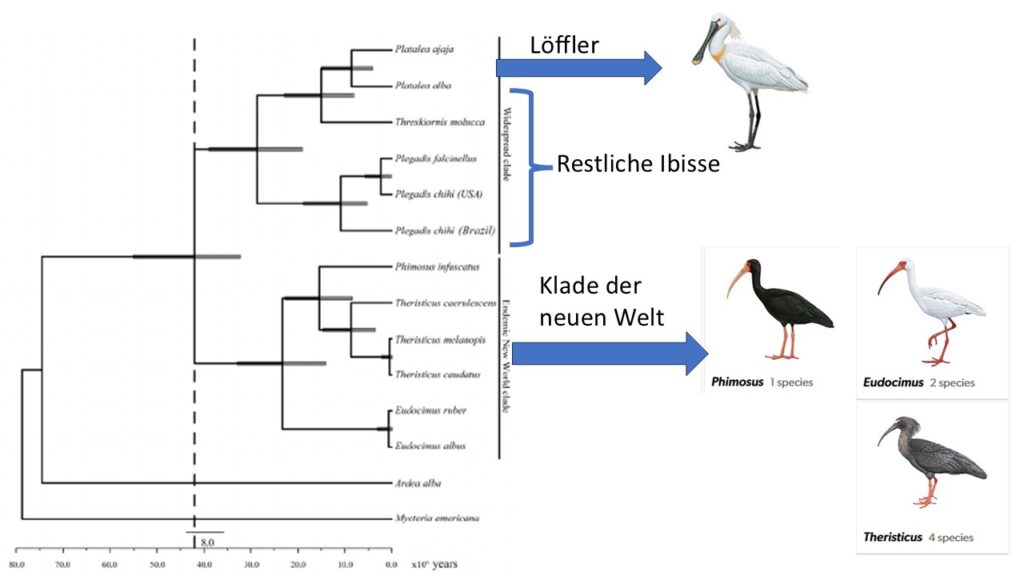
Abb. 3: Phylogenetische Beziehung der Löffler zu den Ibissen.
Die drei neuweltlichen Gattungen, mit einem Schwerpunkt in Mittel- und Südamerika, haben insgesamt 7 Arten, deren bekanntester Vertreter der Scharlachsichler (Eudocimus ruber) sein dürfte. Weitere südamerikanische Arten bzw. Gattungen sind der Langschwanzibis (Gattung Cercibis), der Grünibis (Gattung Mesembrinibis), so wie die drei Arten der Gattung Plegadis, wobei der Braune Sichler (P. falcinellus) weltweit vorkommt und der Brillensichler (P. chihi) hauptsächlich in Nordamerika vorkommt.
Die Gattung Bostrychia kommt mit 5 Arten ausschließlich in Afrika vor. Der Schopfibis (Gattung Lophotibis; L. cristata) nur auf Madagaskar. Die Gattung Nipponia mit einer Art und Pseudibis mit 3, kommen nur in Asien vor. Die eigentlichen Ibisse (Gattung Threskiornis) haben 6 Arten, wobei eine, der Reunion-Ibis (T. solitarius), ausgestorben ist. Die anderen Arten kommen in Asien, Australien und Afrika vor, dessen bekannteste Art der Heilige Ibis (T. aethiopicus) aus Afrika ist. Die Gattung Geronticus hat zwei Arten, den Kahlkopfibis (G. calvus) aus Südafrika und den Waldrapp (G. eremita), der einst in Europa sehr weit verbreitet war. Im 17. Jahrhundert wurden die Waldrappe in Mitteleuropa durch intensive Bejagung sowie den Verlust ihres Lebensraumes ausgerottet. Wild kamen die Waldrappe nur noch in Nordafrika vor.
Nach einem Tiefpunkt Mitte der 1990er Jahre mit nur noch etwa 220 Vögeln in Freiheit ist der Wildbestand seitdem beständig gewachsen. 2019 lebten 700 Vögel wild in Marokko, 325 in der Türkei. Aufgrund verschiedener laufender Wiederansiedlungsprojekte, die das Ziel haben, den Waldrapp erneut als Brutvogel in Europa zu etablieren, gab es 2020/21 wieder ungefähr 340 freilebende Vögel. Der Bestand an wild oder halbwild lebenden Vögeln lag 2020 bei etwa 1400 (Böhm et al. 2020). Darüber hinaus werden etwa 2000 Vögel in Gefangenschaft gehalten.
Die Löffler, Gattung Platalea haben sechs Arten, mit fast weltweiter Verbreitung. In Europa kommt der Eigentliche Löffler (Platalea leucorodia) vor.
Der älteste bekannte Vertreter der Ibisse ist Rhynchaeites messelensis aus dem Eozän, gefunden in der Grube Messel. Weitere Vögel des Eozäns aus China, die Minggangia changgouensis und Ibidopsis hordwelliensis genannt wurden, sind vielleicht ebenfalls Ibisse, obwohl andere Paläontologen sie in die Nähe der Rallen stellen (Peters 1983, Stidham et al. 2005).
Weitere fossile Arten sind in Europa, Asien und Nordamerika beschrieben worden, diese sind jedoch für eine genaue phylogenetische Einordnung zu fragmentarisch belegt (Mayr 2009, 2016, Mayr & Bertelli 2011, M. Wang et al. 2012, Smith et al. 2013). Gerandibis auf dem Miozän Frankreichs gilt als erster Vertreter der Kronengruppe der Ibisse (De Pietri 2013). Im Pliozän treten bereits Vertreter der rezenten Gattungen Threskiornis, Geronticus und Plegadis auf, aus dem Pleistozän sind fossile Überreste rezenter Arten bekannt. Bemerkenswerte Ibisse des späten Pleistozäns und frühen Holozäns sind die flugunfähigen Ibisse von Hawaii (Apteribis) und Jamaica (Xenicibis) (James et al. 1987, Olson & Helen 1991, Olson & Steadman 1977, Tyrberg 2009, Longrich & Olson 2011).
Reiher (Ardeidae)
Reiher sind mit etwa 70 Arten in 20 Gattungen die artenreichste Gruppe der Pelecaniformes und weltweit verbreitet – doch auch hier hat man einen Schwerpunkt in den Tropen, während alle Reiherarten der gemäßigten Breiten Zugvögel sind (Winkler et al. 2015, 2020b, Del Hoyo 2020, Del Hoyo et al. 1992, Kushlan & Hancock 2005, Abb. 4).
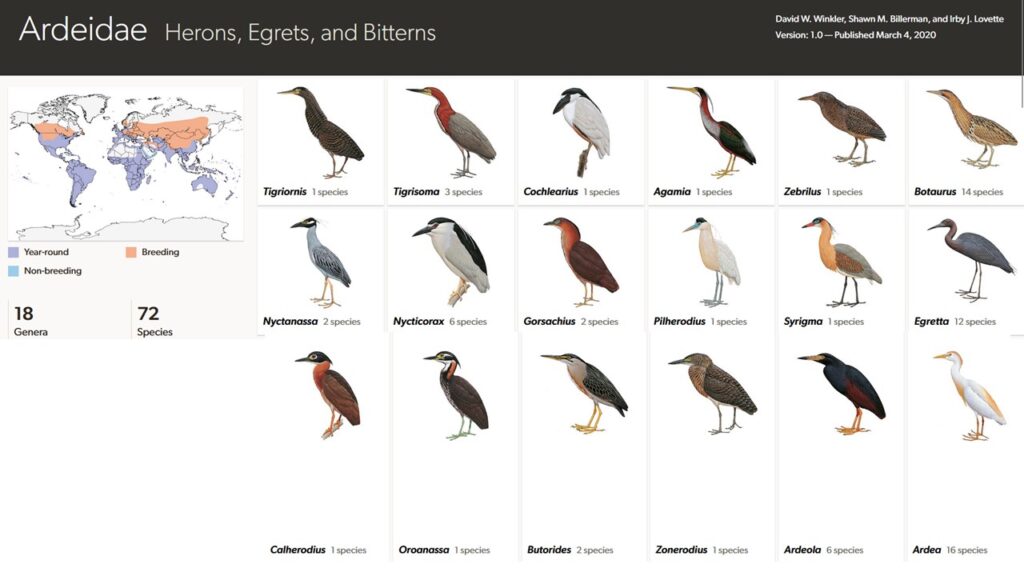
Abb. 4: Reiher
Reiher haben für gewöhnlich einen langen, dolchartigen Schnabel, einen kurzen Schwanz und große, breite Flügel. Der Hals ist S-förmig gebogen, was durch den verlängerten sechsten Halswirbel bedingt ist. Bei fliegenden Reihern ist der gebogene Hals besonders auffällig. Die sogenannten Tagreiher (Unterfamilie Ardeinae) sind meist große Vögel mit ausgeprägt langem Hals und langen Beinen; die Nachtreiher (Unterfamilie Ardeinae; Gattung Nycticorax) und Dommeln (Unterfamilie Botaurinae) sind untersetzter gebaut und wesentlich kurzhalsiger. Zwischen Männchen und Weibchen gibt es nur geringe Größenunterschiede: Männchen sind etwa 2 bis 4 % größer als Weibchen. Ansonsten gibt es keinen auffälligen Geschlechtsdimorphismus, eine Ausnahme bilden allerdings die Zwergdommeln, bei denen das Gefieder von Männchen und Weibchen grundlegend anders gefärbt ist. Viele Arten sind durch verlängerte Schmuckfedern an Kopf, Hals, Brust und/oder Rücken gekennzeichnet. Eine weitere Besonderheit des Reihergefieders sind die Puderdunen. Diese Daunenfedern wachsen ein Leben lang und zerfallen an ihren Spitzen ständig zu einer puderartigen Substanz, die zur Reinigung des Gefieders verwendet wird. Mittelzehe und innere Zehe sind durch eine basale Schwimmhaut miteinander verbunden, ansonsten fehlen Schwimmhäute. Die Beine ermöglichen das Schreiten in tiefem Wasser. Einige Arten haben stark verlängerte Zehen, mit denen sie auf schwimmenden Pflanzenteilen Halt finden. Die Dommeln (Botaurus) können schließlich im Gesträuch und im Röhricht umherklettern. Abgesehen von wenigen Ausnahmen ernähren sich Reiher von Fischen, Amphibien, Reptilien, Wasservögeln, Kleinsäugern, Insekten, Mollusken und Krebstieren. Für gewöhnlich wird die Beute überrascht, indem der Reiher bewegungslos auf der Stelle steht und dann blitzschnell mit dem Schnabel zustößt. Viele Reiher nutzen ihre Flügel beim Beutefang. Sie breiten sie aus, um Schatten zu spenden, in dem potenzielle Beutetiere vermeintlichen Schutz vor der Sonne suchen. Ein anderer Vorteil des Flügelschattens ist, dass er Reflexionen der Wasseroberfläche minimiert und dem Reiher eine bessere Sicht gibt. Viele Reiherarten bedienen sich dieser Technik gelegentlich, der afrikanische Glockenreiher (Egretta ardesiaca) hingegen fast ausschließlich. Mangrovenreiher (Butorides striatus) platzieren gezielt Insekten als Köder auf der Wasseroberfläche, um Fische anzulocken.
Einige Arten haben ein ungewöhnliches Beutespektrum. Die australischen Weißwangenreiher (Egretta novaehollandiae) und afrikanischen Schwarzhalsreiher (Ardea melanocephala) wurden beim Fressen von Aas beobachtet, der Mangrovereiher (Butorides striatus) sogar beim Verzehren von Eicheln. Der Generalist unter den Reihern ist der Kuhreiher (Ardea ibis). Zwar erbeutet auch er gelegentlich die reihertypische Nahrung wie Fische und andere Wassertiere, für gewöhnlich hält er sich aber weit vom Wasser entfernt auf, so dass Insekten zu seiner Hauptbeute geworden sind. Als Kulturfolger sucht er seine Nahrung manchmal auf Müllkippen, ebenso nimmt er große Anteile pflanzlicher Nahrung, die er auch aus Silos stiehlt. Besonders bekannt ist der Kuhreiher aber dafür, große Säugetiere (Rinder, Büffel, Elefanten u. a.) zu begleiten und sie von Hautparasiten zu befreien.
Die meisten Reiher brüten in Kolonien, andere wie die Rohrdommel (Botaurus stellaris) und der Goliathreiher (Ardea goliath) brüten einzeln. Die Nester befinden sich entweder in Bäumen oder im Röhricht. Das Nest wird bei der Zwergdommel (Botaurus minutus) ausschließlich vom Männchen, bei anderen Arten von beiden Partnern gemeinsam gebaut. Das Gelege der Reiher besteht aus ein bis zehn Eiern. Das untere Extrem (ein Ei) tritt beim Bindenreiher (Zonerodius heliosylus) und beim Weißschopfreiher (Tigriornis leucolophus) auf, das obere bei den Zwergdommeln (Botaurus minutus). Bei den allermeisten Arten werden drei bis fünf Eier gelegt. Die Jungen schlüpfen nicht gleichzeitig. Das älteste Junge eines Geleges hat somit einen Wachstumsvorsprung; bei der Fütterung durch die Elternvögel versucht es, die jüngeren Geschwister von der Nahrung zu verdrängen. Dadurch kommt es oft vor, dass die jüngsten Geschwister verhungern, oder aber durch Aggressionen mit Schnabelhieben zu Tode kommen.
Reiher kommen wie erwähnt weltweit vor. Die größte Artenvielfalt herrscht in Süd- und Südostasien (24 Arten), gefolgt von Afrika (21), Süd- und Mittelamerika (20) sowie Australien und Ozeanien (16). Artenärmer sind die Nearktis mit 12 und die Paläarktis mit 9 Arten. Einige Reiher haben sehr weite Verbreitungsgebiete, so findet sich der Silberreiher (Ardea alba), Mangrovenreiher (Butorides striatus, siehe Mendales 2023 für eine alternative Phylogenie) und der Nachtreiher (Nycticorax nycticorax) fast weltweit. Graureiher (Ardea cinerea), Purpurreiher (Ardea purpurea), Rallenreiher (Ardeola ralloides) und Seidenreiher (Egretta garzetta) sind über weite Gebiete in Europa, Afrika und Asien verbreitet. Der in Afrika, Asien und Australien lebende Mittelreiher stellt sich tatsächlich als Artenschwarm heraus und jeder Kontinent hat seine eigene Mittelreiher-Art (Asienmittelreiher, Ardea intermedia; Afrikamittelreiher, Ardea brachyrhyncha und Pazifikmittelreiher, Ardea plumifera). Auch der Kuhreiher hat eine weite Verbreitung. Andere Arten haben eher beschränkte Verbreitungsgebiete: so der Madagaskarreiher (Ardea humbloti) und der Galapagosreiher (Butorides sundevalli), die nur auf den namensgebenden Inseln vorkommen. Der Rotscheitelreiher (Gorsachius goisagi) lebt nur in Japan, der Hainan-Reiher (Gorsachius magnificus) kommt in einem kleinen Gebiet Chinas und Vietnams vor. Der Bindenreiher (Zonerodius heliosylus) lebt nur auf Neuguinea und einige afrikanische Reiherarten haben ebenfalls ein überschaubares Verbreitungsgebiet wie der Dickschnabelreiher (Ardeola idae) und Rotbauchreiher (Ardeola rufiventris). Viele Reiherarten sind Küstenbewohner wie Riffreiher (Egretta sacra), Dreifarbenreiher (Egretta tricolor), Küstenreiher (Egretta gularis) und Meerreiher (Egretta dimorpha). Die größte Reiherart ist der afrikanische Goliathreiher (Ardea goliath) mit 140cm Länge und Flügelspannweite von bis zu 230 cm. Die kleinsten Reiher findet man in der Gattung der Zwergdommeln (Botaurus), von denen manche nur eine Körperlänge von 27 cm erreichen.
Reiher werden in fünf Unterfamilien geteilt (Abb. 5): Die Tagreiher (Ardeinae), die Dommeln (Botaurinae), die Tigerreiher (Tigrisomatinae), sowie die mit jeweils einer Art bestehenden Speerreiher (Agamiinae) und Kahnschnabel (Cochleariinae). Der südamerikanische Kahnschnabel (Cochlearius cochlearius) hat einen für Reiher ungewöhnlichen breiten, dicken Schnabel. Der ebenfalls in Südamerika vorkommende Speerreiher (Agamia agami) hat außergewöhnlich kurze Beine und einen sehr langen Schnabel. Die Unterfamilie der Tigerreiher kommt mit 4 Arten in zwei Gattungen vor. Eine Art lebt in Zentralafrika (Tigriornis leucolophus), die anderen in Südamerika (Gattung Tigrisoma). Diese Vögel haben für Reiher verhältnismäßig dicke Hälse. Die Dommeln kommen weltweit vor und haben 15 Arten, von denen eine, die Schwarzrücken-Zwergdommel (Botaurus novaezelandiae) aus Neuseeland, vor 1900 ausgestorben ist. Allen Arten ist eine gedrungene Gestalt mit einem für Reiher verhältnismäßig kurzen, dicken Hals eigen. Der Schnabel ist verhältnismäßig kurz.

Abb. 5: Phylogenetische Beziehungen der Reiher
Obwohl Fossilfunde von Reihern äußerst selten sind, kann man anhand dieser ablesen, dass Reiher eine sehr alte Vogelgruppe sind: Die Gattung Proardea aus dem Eozän Frankreichs (Mayr 2009) ist der älteste bekannte Reiher, und aus derselben Epoche sind Fußspuren bekannt, die wahrscheinlich von Reihern stammen. Bereits im Miozän lassen sich die Linien der großen Tagreiher sowie der kleinen Nachtreiher erkennen, die teils schon den rezenten Gattungen Ardea, Ardeola, Egretta und Nycticorax zugeordnet werden. Im Pleistozän lebten bereits viele der rezenten Arten. Erst etwa 3.000 v. Chr. ist Ardea bennuides, ein auf der Arabischen Halbinsel verbreiteter Reiher, der die größte bekannte Art ist, die je gelebt hat, ausgestorben (Hoch 1979).
Mehrere fossile Arten sind aus Europa, Afrika und Asien beschrieben worden (Worthy et al. 2013, Mlíkovský 2003, Cheneval et al. 1991, Zelenkov 2011, Mayr 2014, Olson 1985). Ferner ist aus dem Oligozän eine ausgestorbene Familie Xenerodiopidae bekannt, die man für nahe Verwandte der Reiher hält (Rasmussen et al. 1987).
Hammerkopf oder Schattenvogel (Scopidae; Scopus umbretta)
Der Hammerkopf (Scopidae; Scopus umbretta) ist ein bis zu 56cm hoher und 470g schwerer Wasservogel, der nur in Afrika vorkommt und der einzige Vertreter seiner Familie ist. Die Bezeichnung Hammerkopf geht auf die ungewöhnliche Kopfform zurück: Die Art hat einen langgezogenen Schnabel sowie eine nach hinten gerichtete Federhaube. Der im Verhältnis zur Gesamtkörpergröße relativ mächtige Schnabel ist etwa 8 cm lang, seitlich zusammengedrückt und endet in einem kleinen Haken. Er hat Ähnlichkeit mit dem Schnabel des Schuhschnabels, ist aber nicht ganz so gewaltig. Ein Geschlechtsdimorphismus besteht nicht. Die Nahrung besteht zu einem Großteil aus Fischen und Amphibien (Winkler et al. 2015, 2020c, Del Hoyo 2020, Del Hoyo et al. 1992, Abb. 6).
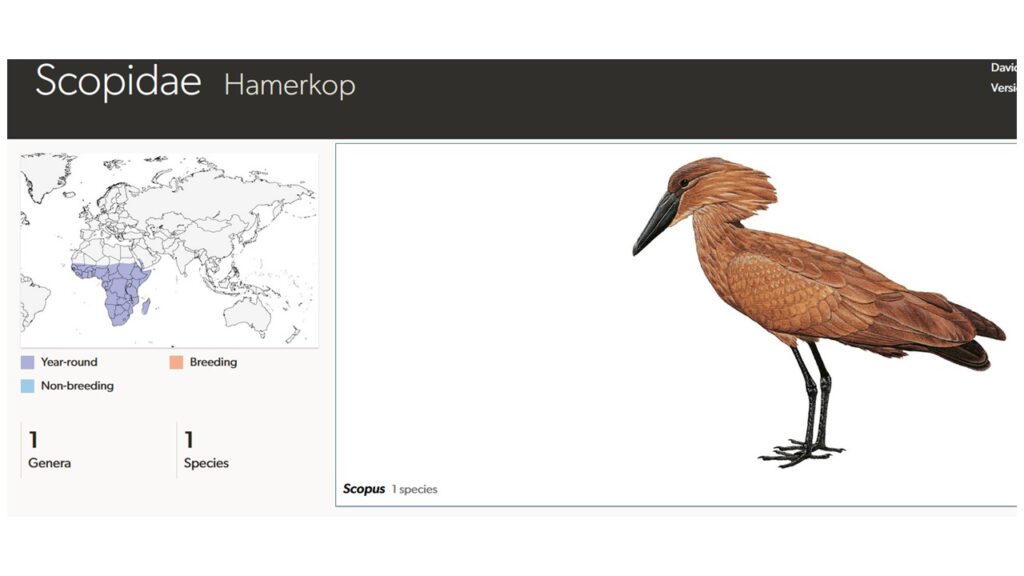
Abb. 6: Hammerkopf
Außergewöhnlich ist das Nest des Hammerkopfes (Abb. 7). Es hat einen Durchmesser von 1,5 m und erreicht das Hundertfache des Eigengewichts des Vogels. Beide Partner bauen es gemeinsam, für gewöhnlich in einem Baum, manchmal auch auf anderen erhöhten Plätzen wie Felsengruppen. Zweige werden aufeinandergehäuft und mit Schlamm aneinandergeklebt. Zunächst bildet das Nest eine Plattform, dann werden die Wände errichtet und schließlich ein Kuppeldach über dem Nest. Der einzige Zugang zu dem riesigen Gebilde befindet sich auf der Unterseite. Ein Paar baut oft mehrere Nester. Diese halten mehrere Jahre und können immer wieder genutzt werden. Das Erweitern und Ausbessern der Nester ist ein beständiger Vorgang, der parallel zur Brut ausgeführt wird. Das Hammerkopfnest dient anderen Tieren als Lebensraum. Kleine Singvögel bauen ihre eigenen Nester auf der Außenseite des Riesennests, und als weitere Untermieter finden sich Honigbienen, Ginsterkatzen, Mangusten, Warane, Schlangen, Nilgänse, Falken, Tauben und Schwarzstörche ein – viele von diesen allerdings nicht gleichzeitig mit einem brütenden Hammerkopfpaar, sondern eher als Bewohner eines verlassenen Nests. Das Gelege besteht aus drei bis sieben Eiern, die beide Partner etwa 30 Tage bebrüten. Beide Partner füttern die Jungen, die nach rund 47 Tagen flügge sind (Wilson et al. 1987, Wilson & Wilson 1986, Martin & Broekhuysen 1961, Kaweesa et al. 2013).

Abb. 7: Nest des Hammerkopfs
Die Bruten sind auffallend häufig erfolglos. So sollen 50 % der Eier unausgebrütet bleiben und 30 bis 40 % der Jungvögel nicht überleben. Hauptursache ist der Einfall von Feinden, vor allem von Waranen, in das Nest. Der Fossilbericht der Familie ist spärlich und beschränkt sich auf eine ausgestorbene Art derselben Gattung aus dem Pliozän Südafrikas (Olson 1984).
Schuhschnabel (Balaenicipitidae; Balaeniceps rex)
Der Schuhschnabel (Balaenicipitidae; Balaeniceps rex) kommt ebenfalls nur in Afrika vor, hat aber ein beschränkteres Verbreitungsgebiet, welches vom Südsudan über Uganda, Westkenia, Westtansania, sowie dem Südosten der Demokratischen Republik Kongo und Nordost-Sambia reicht (Abb. 8). Sein Lebensraum sind Sümpfe und Seeufer, die dicht mit Papyrus oder Schilf bestanden sind. Der Schuhschnabel verdankt seinen Namen der Form seines Schnabels, der außerordentlich lang, aber auch sehr tief und breit wird. Die Schnabellänge liegt bei etwa 22 cm, die Tiefe bei 12cm und die breite bei etwa 10 cm. Einige Individuen haben etwas größere Schnäbel. Der Oberschnabel ähnelt dem der Pelikane; er ist scharf gerandet und trägt eine hakenartige Spitze. Hierdurch kann er schlüpfrige Beutetiere sicher festhalten oder zerteilen. Auch der Kopf ist sehr groß und relativ breit. Der Hals ist hingegen, verglichen mit Störchen und Reihern, relativ kurz. Aufrecht stehend erreicht der Schuhschnabel eine Höhe von 1,20 m. Die langen Beine enden in äußerst langen Zehen, die das Gewicht auf eine große Standfläche verteilen und so ein Einsinken verhindern. Die breiten Flügel ermöglichen einen kräftigen Flug mit ausgedehnten Gleitphasen. Im Flug wird der Kopf wie bei Pelikanen auf die Schultern gelegt, da ein ausgestreckter Hals beim Gewicht des Schnabels nicht vorteilhaft wäre. Das Gefieder ist bläulichgrau, nur der Bauch ist weiß gefärbt. Einen Geschlechtsdimorphismus gibt es nicht. Oft verharrt ein Schuhschnabel lange Zeit bewegungslos. Abgesehen vom Moment des Beutefangs ist sein ganzes Verhalten von langsamen und bedächtigen Bewegungen geprägt. Obwohl er problemlos fliegen kann, nutzt er diese Fähigkeit nur selten. Hauptsächlich ernährt sich der Schuhschnabel von Fischen (Winkler et al. 2015, 2020d, Del Hoyo 2020, Del Hoyo et al. 1992, Fischer 1970, Muller et al. 2015).
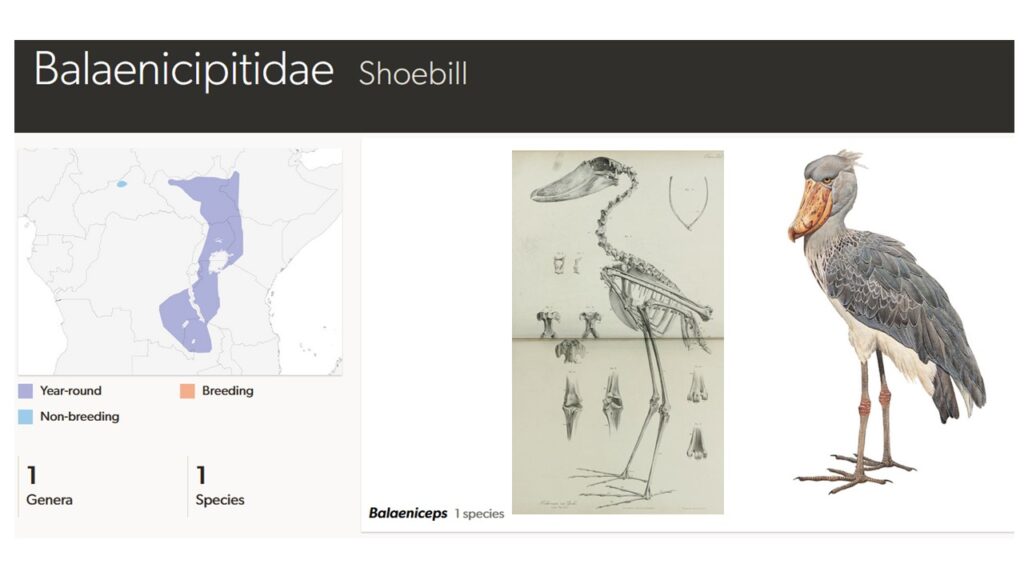
Abb. 8: Schuhschnabel
Das Nest wird inmitten der Sümpfe erbaut, entweder auf festem Untergrund oder auf treibenden Inseln aus Vegetation. Hier wird Pflanzenmaterial zu einem kleinen Hügel angehäuft. Als strikte Einzelgänger verteidigen die Schuhschnabelpaare ein Revier von 2,5 bis 3,8 km² Größe. In das Nest werden ein bis drei Eier gelegt. Beide Partner brüten. Um eine Überhitzung des Geleges zu verhindern, wird es regelmäßig gewendet und mit Wasser übergossen. Die Jungen schlüpfen nach etwa 30 Tagen. Ihr Schnabel ist zunächst noch relativ klein; er beginnt erst in der vierten Lebenswoche zu wachsen. Erst nach 100 Tagen sind die Jungen, die während dieser ganzen Zeit von den Eltern gefüttert werden, flügge. Selbst in diesem Alter haben sie noch einen relativ kleinen Schnabel, der zudem rosa statt gelb ist. Aufgrund des Konkurrenzkampfes unter den Geschwistern um Wasser und Nahrung überlebt oft nur eines der Küken, in der Regel das Erstgeschlüpfte (Del Hoyo et al. 1992, Jasson et al. 2013, Buxton et al. 1978).
Die molekulargenetischen Analysen zeigen, dass der Schuhschnabel von pelikanartigen Vorfahren abstammt, eine Vorstellung, die schon älter ist und vom Ornithologen John Gould im 19. Jh. postuliert wurde. Aufgrund von Untersuchungen des Mittelohrs und der Eischalenstruktur kamen einige Wissenschaftler zu der Überzeugung, Pelikane und Schuhschnabel müssten eng miteinander verwandt sein, während andere behaupteten, alle Gemeinsamkeiten seien das Ergebnis konvergenter Evolution (Mayr 2003, Hagey et al. 2002, Hackett et al. 2008, Jarvis et al. 2014, Prum et al. 2015). Schuhschnabelfossilien aus dem Oligozän und Miozän wurden in Nordafrika entdeckt und der Gattung Goliathia zugeordnet. In Pakistan wurde die Gattung Paludiavis aufgrund einiger Skelettfragmente beschrieben. Diese Funde zeigen, dass schon zu diesem Zeitpunkt Schuhschnäbel recht große Vögel waren und auch außerhalb ihres jetzigen Verbreitungsgebietes anzutreffen waren (Olson 1985, Rasmussen et al. 1987).
Pelikane (Pelecanidae)
Pelikane (Pelecanidae) sind mit 8 Arten in einer Gattung (Pelecanes) mit Ausnahme der Antarktiks auf allen Kontinenten vertreten sind (Winkler et al. 2015, 2020e, Del Hoyo 2020, Del Hoyo et al. 1992, Braun & Urban 1969, Abb. 9).
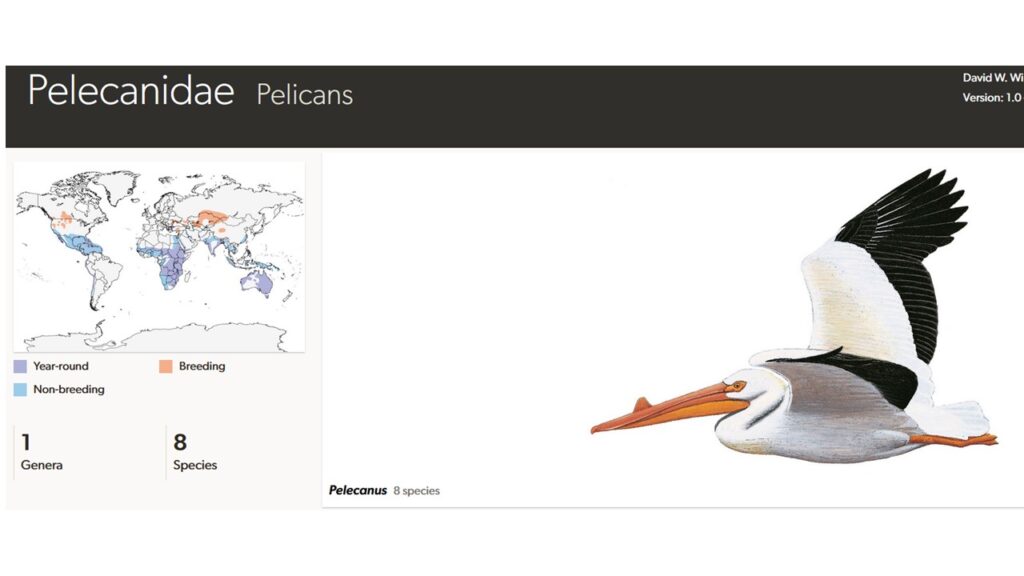
Abb. 9: Pelikane
Pelikane sind sehr große Vögel, die größte Art, der Krauskopfpelikan (P. crispus), hat eine Körperlänge von 1,80 m, eine Flügelspannweite von 3,45 m und ein Gewicht von 13 kg. Als kleinste Art hat der Braune Pelikan (P. occidentalis) eine Spannweite von 2 m und ein Gewicht von 4 kg. Das Skelett macht bei den schwersten Pelikanen nur etwa 7 % des Körpergewichts aus. Das auffälligste Merkmal der Pelikane ist der 25 bis 47 cm lange Schnabel. Der Kehlsack ist bei ihnen extrem vergrößert und mit dem Unterschnabel verbunden, von dem er als außerordentlich dehnbarer Hautsack herabhängt. Dieser Kehlsack, dessen Fassungsvermögen bei den größten Arten bis zu 13 l betragen kann, wird beim Fischfang als Kescher eingesetzt; er wird vom langen, leicht abwärts gebogenen Oberschnabel dicht verschlossen. Die Flügel sind lang und breit, sie sind an sich gute Thermiksegler, ihr Gewicht erschwert aber das Abheben, weswegen Pelikane eine längere Startbahn brauchen. Ist er aber erfolgreich gestartet, ist er ein ausdauernder Flieger. Pelikane können 24 Stunden ohne Pause fliegen und dabei bis zu 500 km zurücklegen; die Fluggeschwindigkeit kann 56 km/h betragen, die Flughöhe über 3000 m. Im Flug biegen Pelikane den Hals zurück, so dass der Kopf zwischen den Schultern liegt und der schwere Schnabel vom Hals abgestützt werden kann. Da die Muskulatur ein ständiges Flügelschlagen nicht erlaubt, wechseln lange Gleitphasen mit Flügelschlägen ab. Hierzu wird die Thermik ausgenutzt, die den Vogel in die Luft trägt und ihm dort ermöglicht, energiesparend zu fliegen.
Die mit Schwimmhäuten versehen Beine liegen weit auseinander und recht weit hinten am Körper. Sie sind geeignet, um beim Schwimmen kräftigen Antrieb zu geben, ermöglichen an Land aber nur eine schwerfällige, watschelnde Fortbewegung. Das Gefieder der Pelikane ist fast immer weiß gefärbt, mit schwarzen Bereichen an den Flügeln. Eine Ausnahme ist der Braune Pelikan (P. occidentalis), dessen Grundfarbe sein Name verrät. Die weißen Pelikane haben manchmal ein rosa oder grau überhauchtes Gefieder; dieser Farbton kommt durch ein Sekret der Bürzeldrüse zustande. Wie kräftig der Farbton ist, steht in einem Zusammenhang mit den regionalen Ernährungsmöglichkeiten.
Die Nahrung der Pelikane besteht fast ausschließlich aus Fischen. Es gibt nur wenige Ausnahmen: So hat man den Nashornpelikan gelegentlich beim Fressen von Schwanzlurchen und Flusskrebsen beobachtet, und auch der Brillenpelikan (P. conspicillatus) ernährt sich nebenher von Krebstieren. In seltenen Fällen verschlingen Pelikane auch ganze Vögel. Ein Pelikan frisst für gewöhnlich zehn Prozent seines Körpergewichts am Tag. Dies sind beim Rosapelikan (P. onocrotalus) etwa 1,2 kg. Rechnet man dies hoch, so verspeist die gesamte Pelikanpopulation des afrikanischen Nakurusees 12.000 kg Fisch am Tag bzw. 4380 Tonnen Fisch im Jahr. Pelikane brüten in Kolonien, wobei die Bodenbrüter größere und dichtere Kolonien bilden als die Baumbrüter. Oft entstehen gemischte Kolonien: So brüten im Donaudelta Rosa- und Krauskopfpelikane oft gemeinsam; die baumbrütenden Arten nisten neben Störchen und Kormoranen, die Braunpelikane neben Guanotölpeln und Guanokormoranen.
Zur Balz nehmen die nackten Hautpartien im Gesicht leuchtende Farben ein. Die männlichen Pelikane vollführen ein Balzritual, das sich von Art zu Art unterscheidet, oft aber ein Emporrecken von Kopf und Schnabel und ein ballonartiges Aufblasen des Hautsacks am Unterschnabel beinhaltet. Nachdem sich das Paar gefunden hat, sucht das Weibchen (beim Braunpelikan das Männchen) einen Nistplatz. Sodann beginnt der Nestbau, der wiederum von Art zu Art sehr unterschiedlich ist. Die bodenbrütenden Arten legen oft nur eine Mulde aus, die sie mit keinem bis wenig Material wie Gräsern und Zweigen ausfüllen. Natürlich sind die Nester der baumbrütenden Arten aufwändiger gestaltet. Der Graupelikan (P. philippensis) brütet vor allem auf Mangobäumen, Feigen, Palmyrapalmen oder Kokospalmen. Baumaterial wird vom Männchen in seinem Schnabelsack herbeigeschafft. Das Nest besteht aus Zweigen und wird mit Gräsern oder verfaulenden Wasserpflanzen ausgelegt; es hat einen Durchmesser von etwa 75 cm und eine Höhe von 30 cm. Die Stabilität des Nestes ist oft gering, so dass jedes Jahr ein neues Nest gebaut werden muss.
Meistens werden zwei Eier gelegt, Gelege mit nur einem und bis zu sechs Eiern kommen vor. Beide Geschlechter brüten, die Gesamtbrutdauer beträgt 30 bis 36 Tage. Die Jungvögel sind anfangs nackt, entwickeln aber schon nach wenigen Tagen das je nach Art weiße oder braune Daunenkleid. Im Alter von acht Wochen ist das Daunenkleid durch das Jugendgefieder ersetzt. Anfangs werden die Jungen mit einem ausgewürgten Nahrungsbrei gefüttert. Selten kommen alle Jungvögel durch. Oft erweist sich das zuerst geschlüpfte Junge als das stärkere, das seine Geschwister aus dem Nest drängt oder durch alleinige Beanspruchung der Nahrung die anderen verhungern lässt. Im Alter von 70 bis 85 Tagen werden die Jungen selbständig und verlassen die Eltern entweder sofort oder nach einer Frist von bis zu 20 Tagen, in denen sie noch gemeinsam mit den Elternvögeln unterwegs sind.
Pelikane bewohnen tropische, subtropische und gemäßigte Zonen. In Europa befinden sich Vorkommen auf dem Balkan, wobei die Kolonien des Rosa- und Krauskopfpelikans (P. onocrotalus; P. crispus) im Donaudelta die bekanntesten sind. Daneben bestehen noch Vorkommen dieser beiden Arten am Prespasee und an der Ostküste des Asowschen Meeres. Der Krauskopfpelikan kommt in einigen Kolonien am Unterlauf der Wolga sowie an der Nordküste des Kaspischen Meeres vor. Diese beiden Arten sowie den Graupelikan (P. philippensis) findet man auch in West- und Zentralasien. Letzteren zusätzlich in Südasien. Afrika ist die Heimat des Rötelpelikans (P. rufescens), der dort tropische und subtropische Regionen bewohnt; weiter gespannt sind die Brut- und Winterquartiere des Rosapelikans (P. onocrotalus), dessen Verbreitung von der Sahelzone bis Südafrika reicht. In Australien und Tasmanien lebt der Brillenpelikan (P. conspicillatus), der außerhalb der Brutzeit regelmäßig auf Neuguinea, den Salomonen und den Kleinen Sunda-Inseln anzutreffen ist. Der Nashornpelikan (P. erythrorhynchos) brütet im Mittleren Westen Nordamerikas, nordwärts bis ins südliche Kanada; er überwintert an den Küsten Nord- und Mittelamerikas. Die Küsten des amerikanischen Doppelkontinents sind die Heimat des Braunpelikans (P. occidentalis). Der Chile-Pelikan (P. thagus) lebt an der Pazifikküste von Peru und Chile.
Ein früher möglicher Vertreter der Pelikane ist Protopelicanus cuvieri aus dem Eozän, dessen Einordnung in die Pelikane jedoch umstritten ist und einige Autoren sehen eine Verwandtschaft mit den Tölpeln oder den ausgestorbenen Pseuozahnvögeln (Harrison 1979, Olson 1985a). Der älteste zweifelsfreie Pelikan stammt aus dem Miozän Frankreichs und heißt Pelecanus gracilis (manchmal auch in einer eigenen Gattung Miopelecanus geführt). Kurz später tauchte Pelecanus intermedius auf, ein in Deutschland oft gefundener fossiler Pelikan, sowie der kleine Pelecanus tirarensis in Australien. Im Pliozän gab es noch weit mehr Arten, darunter mit dem Nashornpelikan (P. erythrorhynchos) auch bereits eine rezente Art, und im Pleistozän finden sich Überreste mehrerer rezenter Arten. Das plötzliche Auftauchen mehrerer Arten im Fossilbericht Europas deutet darauf hin, dass die Pelikane woanders entstanden sind. Aufgrund der verwandtschaftlichen Nähe zum Schuhschnabel kann ein möglicher Ursprung in Afrika gesehen werden (Lydekker 1891, El Adli et al. 2021, Miller 1966, Rich & van Tets 1981, Wetmore 1933, Widhalm 1886, Noriega et al. 2023, Louchart et al. 2011).
Veränderungen in der Artenzusammensetzung bei Seevögeln
In den vorherigen Episoden haben wir eine Reihe an Vogelgruppen kennengelernt, die an aquatische Lebensräume gebunden sind. Neben den in dieser und den vorherigen beiden Episoden vorgestellten Aequornithes zählen Gänsevögel und die Tropikvögel, aber auch die Charadriiformes, also Alkenvögel, Möwen und Regenpfeierartige zu den hauptsächlich aquatischen Vögeln. Viele dieser Vögel sind auch auf die offenen Ozeane angepasst.
Die Zusammensetzung der marinen Avifaunen des frühen Känozoikums unterscheidet sich deutlich von der der heutigen, wo die artenreichsten und am weitesten verbreiteten Gruppen von Meeresvögeln Enten und Gänse, die Sturmschwalben- und Sturmvögel, Kormorane, Möwen, Skuas und Alke sind.
Viele dieser Gruppen scheinen im Verlauf der Evolutionsgeschichte vergleichsweise spät in marine Lebensräume eingedrungen zu sein. Verschiedene artenarme Meeresvögelgruppen, kommen hauptsächlich in der südlichen Hemisphäre vor, hatten aber in der Vergangenheit eine größere Verbreitung, Artenvielfalt und verschiedene ökologische Nischen, wie z. B. Tropikvögel, Fregattvögel, Tölpel und Albatrosse.
Fregattvögel, die heute ausschließlich in marinen Lebensräumen vorkommen, kamen im frühen Eozän auch in Süßwassergebieten vor, und im Gegensatz zu den heutigen, stark pelagisch lebenden Tropikvögeln hielten sich die fossilen Formen dieser Gruppe im frühen Eozän in küstennahen Lebensräumen auf.
Mehrere Seevogeltaxa wie die Pinguine, die Röhrennasen und die Tropikvögel haben eine lange Evolutionsgeschichte in marinen Umgebungen, während von den großen marinen Möwen und Raubmöwen keine Aufzeichnungen aus Ablagerungen vor dem Pliozän existieren.
In der letztgenannten Epoche scheinen sich einige der dramatischsten Veränderungen in den globalen Seevogelgemeinschaften vollzogen zu haben. Insbesondere in der nördlichen Hemisphäre kam es während des Pliozäns zu einem erheblichen Verlust an Vielfalt bei den Seevögeln (Warheit 1992, 2002, Olson & Rasmussen 2001, Olson & Hearty 2003, Smith 2011), und zu den Gruppen, die lokal oder global ausgestorben sind, gehören Pelagornithidae, Albatrosse im Nordatlantik und die flugunfähigen Mancallinae im Nordpazifik.
Die ozeanischen Zirkulationssysteme waren während des Känozoikums erheblichen Veränderungen unterworfen, die auf wichtige geografische Ereignisse wie die Öffnung der Drake-Passage zwischen Südamerika und der Antarktis zurückzuführen sind, die im späten Eozän zur Bildung des zirkumantarktischen Stroms führte (Scher & Martin 2006). Der Golfstrom, eine der wichtigsten ozeanischen Strömungen der nördlichen Hemisphäre, scheint sich im Pliozän gebildet zu haben, und seine Entstehung wurde lange Zeit mit der Entstehung des panamaischen Isthmus in Verbindung gebracht, der heute den Atlantik vom Pazifik trennt (Haug & Tiedemann 1998). In jüngster Zeit wurde jedoch eine Entstehung des Isthmus in der Mitte des Miozäns vorgeschlagen (Montes et al. 2015), was besser mit der Tatsache übereinstimmt, dass die miozänen Seevogelgemeinschaften des Atlantiks und des Pazifiks bereits sehr unterschiedlich waren. Weder Plotopteridae noch Mancallinae kommen außerhalb der nordpazifischen Region vor, was auf das Vorhandensein physischer oder ökologischer Ausbreitungsbarrieren hindeutet, die im Falle der Plotopteridae bereits vor der Entstehung der panamaischen Landenge im mittleren Miozän bestanden haben müssen.
Die Faktoren, die die Evolution der Seevögel beeinflusst haben, sind noch nicht genau bekannt, und neben den Veränderungen der ozeanischen Zirkulationssysteme und der Meeresproduktivität könnte sich auch der Raubbau an den Nistplätzen auf die Seevogelgemeinschaften ausgewirkt haben. Selbst die meisten pelagischen Vögel müssen an Land gehen, um ihre Küken aufzuziehen. Aufgrund der langen Brut- und Nistzeiten und der geringen Anzahl von Nachkommen sind Seevögel an ihren Nistplätzen besonders anfällig für Raubtiere. Viele der heute lebenden Seevögel brüten daher in abgelegenen oder unzugänglichen Gebieten, wie z. B. auf ozeanischen Inseln, an steilen Klippen oder in Regionen mit ungünstigen klimatischen Bedingungen in hohen Breitengraden, wo die Konkurrenz um Brutplätze groß sein kann. Es ist bekannt, dass eingeschleppte Raubsäugetiere wie Katzen oder Ratten verheerende Auswirkungen auf bestehende Meeresvogelfaunen haben, und das späte Auftreten von Skuas und Möwen im Känozoikum, die wichtige Eier- und Nestlingsräuber in bestehenden Seevogelkolonien sind, könnte den Wettbewerb um geeignete Brutplätze noch verschärft haben. Die Veränderungen des Meeresspiegels während des Känozoikums führten schließlich zu einer unterschiedlichen Verfügbarkeit von vorgelagerten Inseln als Brutplätze. Das Verschwinden sicherer Brutplätze auf vorgelagerten Inseln wurde beispielsweise als Ursache für das lokale Aussterben der Plotopteridae angesehen (Goedert & Cornish 2002), und pliozäne Schwankungen des Meeresspiegels (Dwyer & Chandler 2009) könnten sich ebenfalls auf einige ökologisch weniger flexible Seevogeltaxa ausgewirkt haben, wie etwa die flugunfähigen Mancallinae Alken und die Riesenpelagornithiden.
Die letzte Großklade der Vögel, die behandelt werden müssen sind die Telluraves, die Landvögel. Für die restlichen 9 Kladen der Neoaves, sowie für die Urkriefervögel und Hühner- und Gänsevögel haben wir 10 Episoden gebraucht. Aber die Telluraves machen die mit Abstand größte Klade innerhalb der Vögel aus. Hier finden sich Greifvögel, Spechte, Eulen, Papageien und natürlich die Sperlingsvögel. Sicher ist, dass wir für diese Gruppe wohl ebenfalls viele Episoden machen werden.
Literatur
Böhm, C., Bowden, C., Seddon, P. et al. (2020): The northern bald ibis Geronticus eremita: history, current status and future perspectives. Cambridge University Press, doi:10.1017/S0030605320000198
Braun, E. L.; Kimball, R. T. (2021): Data types and the phylogeny of Neoaves. Birds. 2 (1): 1–22.
Brown, L. H., Urban, E. K. (1969): The breeding biology of the Great White Pelican Pelecanus onocrotalus roseus at Lake Shala, Ethiopia. Ibis 111, S. 199–237
Buxton, L.; Slater, J.; Brown, L. H. (1978): The breeding behaviour of the shoebill or whale-headed stork Balaeniceps rex in the Bangweulu Swamps, Zambia. African Journal of Ecology. 16 (3): 201–220.
Cheneval, J., Ginsburg, L., Mourer-Chauviré, C., Ratanasthien, B. (1991): The Miocene avifauna of the Li Mae Long locality, Thailand: Systematics and paleoecology. Journal of Southeast Asian Earth Sciences 6, 117–126.
Chesser, R. T., Yeung, C. K. L., Yao, C.-T., Tian, X.-H., Li, S.-H (2010): Molecular phylogeny of the spoonbills (Aves: Threskiornithidae) based on mitochondrial DNA. Zootaxa, V. 2603
Del Hoyo, J. (2020): All the Birds of the World. Lynx Nature Books
Del Hoyo, J.; Elliot, A.; Sargatal, J., eds. (1992): Handbook of the Birds of the World, volume 1, Ostrich to Dodos, Lynx Edicions, Barcelona.
De Pietri, V.L. (2013): Interrelationships of the Threskiornithidae and the phylogenetic position of the Miocene ibis “Plegadis” paganus from the Saint-Gérand-le-Puy area in central France. The Ibis 155, 544–560.
Dwyer, G.S., Chandler, M.A. (2009): Mid-Pliocene sea level and continental ice volume based on coupled benthic Mg/Ca palaeotemperatures and oxygen isotopes. Philosophical Transactions of the Royal Society A: Mathematical, Physical and Engineering Sciences 367, 157–168.
El Adli, J. J.; Wilson M., Jeffrey A.; Antar, M. S. M.; Gingerich, P. D. (2021): The earliest recorded fossil pelican, recovered from the late Eocene of Wadi Al-Hitan, Egypt. Journal of Vertebrate Paleontology. 41 (1): e1903910.
Fischer, W. (1970): Der Schuhschnabel. Die neue Brehm-Bücherei
Goedert, J.L., Cornish, J. (2002): A preliminary report on the diversity and stratigraphic distribution of the Plotopteridae (Pelecaniformes) in Paleogene rocks of Washington State, USA. In: Z.-H. Zhou & F.-Z. Zhang (eds.), Proceedings of the 5th Symposium of the Society of Avian Paleontology and Evolution, 1-4 June 2000. Beijing: Science Press, pp. 63–76.
Hackett, S. J. et al. (2008): A phylogenomic study of birds reveals their evolutionary history. Science 320: 1763–1768.
Hagey, J. R.; Schteingart, C. D.; Ton-Nu, H.-T., Hofmann, A. F. (2002): A novel primary bile acid in the Shoebill stork and herons and its phylogenetic significance. Journal of Lipid Research. 43 (5): 685–90. doi:10.1016/
Harrison, C. J. O. (1979): The Upper Eocene birds of the Paris basin; a brief re-appraisal. Tertiary Research 2, S. 105–109
Haug, G.H., Tiedemann, R. (1998): Effect of the formation of the Isthmus of Panama on Atlantic Ocean thermohaline circulation. Nature 393, 673–676.
Hoch, E. (1979): Reflections on Prehistoric Life at Umm an-Nar (Trucial Oman) Based on Faunal Remains from the Third Millennium B.C. South Asian Archaeology S. 589–638
Houde, P. et al. (2019): Phylogenetic Signal of Indels and the Neoavian Radiation. Diversity. 11 (7): 108.
James, H. F., et al. (1987): Radiocarbon Dates on Bones of Extinct Birds from Hawaii. doi:10.1073/pnas.84.8.2350;84;2350-2354 PNAS 2 8.
James, H. F., Olson, S. L (1991): Descriptions Of Thirty-Two New Species Of Birds From The Hawaiian Islands: Part I. Non-Passeriformes. Ornithological Monographs No. 45 Published By The American Ornithologists’ Union Washington, D.C. University of California. Berkeley, California
Jarvis, E. D.; et al. (2014): Whole-genome analyses resolve early branches in the tree of life of modern birds. Science. 346 (6215): 1320–1331
Jasson, J.; Cuthbert, N.; Woo, L.; Charles, M. (2013): Observations on nesting of shoebill Balaeniceps rex and wattled crane Bugeranus carunculatus in Malagarasi wetlands, western Tanzania. African Journal of Ecology. 51 (1): 184–187.
Kaweesa, S.; Jonkvorst, R.; Katebaka, R.; Ssemmanda, R.; Pomeroy, D.; Brouwer, J. (2013): Is the Hamerkop Scopus umbretta a neocolonist or an opportunist nester? Scopus. 32: 35–38.
Kimball, RT et al. (2013): Identifying localized biases in large datasets: A case study using the Avian Tree of Life. Mol Phylogenet Evol. 69 (3): 1021–32.
Kuhl, H. et al. (2021): An unbiased molecular approach using 3’UTRs resolves the avian family-level tree of life. Molecular Biology and Evolution. 38: 108–127.
Kuramoto T, Nishihara H, Watanabe M, Okada N (2015): Determining the position of storks on the phylogenetic tree of waterbirds by retroposon insertion analysis, Genome Biol. Evol. 7, 3180-89.
Kushlan, J. A., Hancock, J. A. (2005): Herons. Oxford University Press
Longrich, N. R.; Olson, S. L. (2011): The bizarre wing of the Jamaican flightless ibis Xenicibis xympithecus: a unique vertebrate adaptation. Proceedings of the Royal Society B: Biological Sciences. 278 (1716): 2333–2337.
Louchart, A.; Tourment, N.; Carrier, J. (2011): The earliest known pelican reveals 30 million years of evolutionary stasis in beak morphology. Journal of Ornithology 152(1), 15–20. doi:10.1007/s10336-010-0537-5
Lydekker, R. (1891): Catalogue of the Fossil Birds in the British Museum (Natural History). London, United Kingdom: British Museum. pp. 37–45.
Martin, J.; Broekhuysen, G. J. (1961): Some records of birds using the nests of others. Ostrich. 32 (3): 104–106.
Mayr, H. (2003): The phylogenetic affinities of the Shoebill (Balaeniceps rex). Journal of Ornithology 144, S. 157–175.
Mayr, G. (2009): Paleogene Fossil Birds. Heidelberg: Springer
Mayr, G. (2014): On the middle Miocene avifauna of Maboko Island, Kenya. Geobios 47, 133–146.
Mayr, G. (2016): Avian Evolution: The Fossil Record of Birds and its Paleobiological Significance. TOPA Topics in Paleobiology.
Mayr, G., Bertelli, S. (2011): A record of Rhynchaeites (Aves, Threskiornithidae) from the early Eocene Fur Formation of Denmark, and the affinities of the alleged parrot Mopsitta. Palaeobiodiversity and Palaeoenvironments 91, 229–236.
Mendales, E. Z. (2023): Ultraconserved elements resolve the phylogeny of a globally distributed genus, Butorides (Aves: Ardeidae). MS Thesis, San Francisco State University, doi: 10.46569/hd76s649j
Miller, A.H. (1966): The Fossil Pelicans of Australia. Memoirs of the Queensland Museum. 14: 181–90.
Mlíkovský, J. (2003): Early Miocene birds of Djebel Zelten, Lybia. Casopis Národníhomuzea, Rada prírodovedná 172, 114–120.
Montes, C., Cardona, A., Jaramillo, C., Pardo, A., Silva, J.C., Valencia, V., Ayala, C., Pérez-Angel, L.C., Rodriguez-Parra, L.A., Ramirez, V., Niño, H. (2015): Middle Miocene closure of the Central American Seaway. Science 348, 226–229.
Mullers, R. HE; Amar, A. (2015): Shoebill Balaeniceps rex foraging behaviour in the Bangweulu Wetlands, Zambia. Ostrich. 86 (1–2): 113–118.
Noriega, J. I.; Cenizo, M.; Brandoni, D.; Pérez, L. M.; Tineo, D. E.; Diederle, J. M.; Bona, P. (2023): A new pelican (Aves: Pelecanidae) from the Upper Miocene of Argentina: new clues about the origin of the New World lineages. Journal of Vertebrate Paleontology. 42 (5): e2202702.
Olson, S.L. (1984): A hamerkop from the early Pliocene of south Africa (Aves: Scopidae). Proceedings of the Biological Society of Washington 97, 736–740.
Olson, S.L. (1985): The fossil record of birds. In: D.S. Farner, J.R. King, & K.C. Parkes (eds.), Avian Biology, vol. 8. New York: Academic Press, pp. 79–238.
Olson, S. L. (1985a): A selective synopsis of the fossil record of birds. In: D. Farner, J. R. King, K. Parkes: Avian Biology 8. Academic Press, New York 1985
Olson, S.L., Hearty, P.J. (2003): Probable extirpation of a breeding colony of Short-tailed Albatross (Phoebastria albatrus) on Bermuda by Pleistocene sea-level rise. Proceedings of the National Academy of Sciences USA 100, 12825–12829.
Olson, S. L.; James, H. (1991): Descriptions of Thirty-Two New Species of Birds from the Hawaiian Islands Part I. Non-Passeriformes“. Ornithological Monographs. 7 (45): 1–88.
Olson, S.L., Rasmussen, P.C. (2001): Miocene and Pliocene birds from the Lee Creek Mine, North Carolina. Smithsonian Contributions to Paleobiology 90, 233–365.
Olson, S. L; Steadman, D. W. (1977): A new genus of flightless ibis (Threskiornithidae) and other fossil birds from cave deposits in Jamaica. Proceedings of the Biological Society of Washington. 90 (2): 447–457.
Peters, D. S. (1983): Die „Schnepfenralle“ Rhynchaeites messelensis Wittich 1898 ist ein Ibis. Journal of Ornithology 124, Nr. 1, S. 1–27
Prum, R. O. et al. (2015): A comprehensive phylogeny of birds (Aves) using targeted next-generation DNA sequencing. Nature. 526 (7574): 569–573.
Ramirez, J. L., Miyaki, C. Y., Del Lama, S. N. (2013): Molecular phylogeny of Threskiornithidae (Aves: Pelecaniformes) based on nuclear and mitochondrial DNA. Genetics and molecular research 12, Nr. 3, S. 2740–2750
Rasmussen, D.T., Olson, S.L., Simons, E.L. (1987): Fossil birds from the Oligocene Jebel Qatrani Formation, Fayum Province, Egypt. Smithsonian Contributions to Paleobiology 62, 1–20.
Rich, P.V.; van Tets, J. (1981): The Fossil Pelicans of Australia. Records of the South Australian Museum (Adelaide). 18 (12): 235–64.
Sangster, G; Braun, EL; Johansson, US; Kimball, RT.; Mayr, G; Suh, A (2022): Phylogenetic definitions for 25 higher-level clade names of birds. Avian Research. 13: 100027
Sangster G, Mayr G (2021): Feraequornithes: a name for the clade formed by Procellariiformes, Sphenisciformes, Ciconiiformes, Suliformes, and Pelecaniformes (Aves), Vertebr. Zool. 71, 49-53.
Scher, H.D., Martin, E.E. (2006): Timing and climatic consequences of the opening of Drake Passage. Science 312, 428–430.
Smith, N.A. (2011): Taxonomic revision and phylogenetic analysis of the flightless Mancallinae (Aves, Pan-Alcidae). Zookeys 91, 1–116.
Smith, N.D., Grande, L., Clarke, J.A. (2013): A new species of Threskiornithidae-like bird (Aves, Ciconiiformes) from the Green River Formation (Eocene) of Wyoming. Journal of Vertebrate Paleontology 33, 363–381.
Stidham, T. A. et al. (2005): An ibis-like bird (Aves: Threskiornithidae) from the late middle Eocene of Myanmar. Contributions from the museum of paleontology of the University of Michigan 31, Nr. 7, S. 179–184.
Suh, A. (2016): The phylogenomic forest of bird trees contains a hard polytomy at the root of Neoaves. Zoologica Scripta. 45: 50–62.
Tyrberg, T. (2009): Holocene avian extinctions. In Turvey, Sam (ed.). Holocene Extinctions. Oxford University Press. pp. 63–106.
Van Tuinen, M., Butvill, D. B., Kirsch, J. A. W., Hedges, S. B. (2001): Convergence and Divergence in the Evolution of Aquatic Birds. Proceedings of the Royal Society: Biological Sciences 268, Nr. 1474, S. 1345–1350.
Warheit, K.I. (1992): A review of the fossil seabirds from the Tertiary of the North Pacific: Plate tectonics, paleoceanography, and faunal change. Paleobiology 18, 401–424.
Warheit, K.I. (2002): The seabird fossil record and the role of paleontology in understanding seabird community structure. In: E.A. Schreiber & J. Burger (eds.), Biology of Marine Birds. Boca Raton, FL: CRC Marine Biology Series, pp. 17–55.
Wang, M., Mayr, G., Zhang, J., Zhou, Z. (2012): New bird remains from the Middle Eocene of Guangdong, China. Acta Palaeontologica Polonica 57, 519–526.
Wetmore, A. (1933): Pliocene Bird Remains from Idaho. Smithsonian Miscellaneous Collections. 87 (20): 1–12.
Widhalm, J. (1886): Die Fossilen Vogel-Knochen der Odessaer-Steppen-Kalk-Steinbrüche an der Neuen Slobodka bei Odessa. Schriften der Neurussische Gesellschaft der Naturforscher zu Odessa (in German). 10: 3–9
Wilson, R.; Wilson, M. (1986): Nest building by the Hamerkop Scopus umberetta. Ostrich. 57 (4): 224–232.
Wilson, R. T.; Wilson, M. P.; Durkin, J. W. (1987): Aspects of the reproductive ecology of the Hamerkop Scopus umbretta in central Mali. Ibis. 129 (2): 382–388.
Winkler, D. W., Billerman, S. M., Lovette, I. J. (2015): Bird Families of the World: A Guide to the Spectacular Diversity of Birds. Lynx Edicions, Barcelona.
Winkler, D. W., S. M. Billerman, and I. J. Lovette (2020a). Ibises and Spoonbills (Threskiornithidae), version 1.0. In Birds of the World (S. M. Billerman, B. K. Keeney, P. G. Rodewald, and T. S. Schulenberg, Editors). Cornell Lab of Ornithology, Ithaca, NY, USA. https://doi.org/10.2173/bow.thresk1.01
Winkler, D. W., S. M. Billerman, and I. J. Lovette (2020b). Herons, Egrets, and Bitterns (Ardeidae), version 1.0. In Birds of the World (S. M. Billerman, B. K. Keeney, P. G. Rodewald, and T. S. Schulenberg, Editors). Cornell Lab of Ornithology, Ithaca, NY, USA. https://doi.org/10.2173/bow.ardeid1.01
Winkler, D. W., S. M. Billerman, and I. J. Lovette (2020c). Hamerkop (Scopidae), version 1.0. In Birds of the World (S. M. Billerman, B. K. Keeney, P. G. Rodewald, and T. S. Schulenberg, Editors). Cornell Lab of Ornithology, Ithaca, NY, USA. https://doi.org/10.2173/bow.scopid1.01
Winkler, D. W., S. M. Billerman, and I. J. Lovette (2020d). Shoebill (Balaenicipitidae), version 1.0. In Birds of the World (S. M. Billerman, B. K. Keeney, P. G. Rodewald, and T. S. Schulenberg, Editors). Cornell Lab of Ornithology, Ithaca, NY, USA. https://doi.org/10.2173/bow.balaen1.01
Winkler, D. W., S. M. Billerman, and I. J. Lovette (2020e). Pelicans (Pelecanidae), version 1.0. In Birds of the World (S. M. Billerman, B. K. Keeney, P. G. Rodewald, and T. S. Schulenberg, Editors). Cornell Lab of Ornithology, Ithaca, NY, USA. https://doi.org/10.2173/bow.peleca1.01
Worthy, T.H., Worthy, J.P., Tennyson, A.J.D., Scofield, R.P. (2013): A bittern (Aves: Ardeidae) from the Early Miocene of New Zealand. Paleontological Journal 47, 1331–1343.
Wu, S. et al. (2024): Genomes, fossils, and the concurrent rise of modern birds and flowering plants in the Late Cretaceous. Proceedings of the National Academy of Sciences. 121 (8).
Zelenkov, N.V. (2011): Ardea sytchevskayae sp. nov., a new heron species (Aves: Ardeidae) from the Middle Miocene of Mongolia.
