
Missing links?
Kreationisten sind verliebt in Fossilien. Sie wissen zwar nicht viel davon, denn meist referieren sie aus Kinderbüchern über Dinosaurier, reden aber umso mehr wirres Zeug darüber. Schließlich stacheln sie sich untereinander auf, gebetsmühlenartig zu widerholen, es gebe nichts als Lücken im Fossilbericht. Sie stellen die Behauptung auf, dass wenn Evolution wahr wäre, müssten es Millionen und Abermillionen von sogenannten Übergangsformen geben. Und diese Übergangsformen müsste man im Fossilbericht wiederfinden. Diese gäbe es jedoch nicht, behaupten sie dann aber. Wenn man ihnen dann einige Übergangsformen zeigt, z. B. Archaeopteryx, dann sagen sie, dass Archaeopteryx nicht den Übergang von Dinosauriern zu Vögeln zeige, sondern einfach nur ein fertiger Vogel sei. Bei genauerer Betrachtung wird man jedoch feststellen, dass Kreationisten eine sehr obskure Vorstellung dessen haben, was eine Übergangsform ist und ihnen jegliche Kenntnis der Anatomie, Systematik, ja gar der Biologie und Wissenschaft überhaupt fehlt. Schauen wir uns mal einige ihrer Vorstellungen an:
Video dazu:

![]()
Abb.1 Vorstellungen der Kreationisten, wie Übergangsformen aussehen sollen
Wir sehen also: gerne nehmen Kreationisten Körperteile von heute lebenden Tieren und fügen sie zu abstrusen Monstern zusammen, die an Peinlichkeiten nur von Filmen wie Angriff der Killertomaten überboten werden können. Da wird einer Ente ein Krokodilkopf aufgesetzt und Croco-Duck getauft, weil Evolutionsbiologen nun mal festgestellt haben, dass Vögel und Krokodile zusammen die Gruppe der Archosaurier bilden und ein Übergang zwischen beiden Gruppen könne nur diese Crocoduck sein – da kann Archaeopteryx natürlich nicht mithalten.
Wir sollten aber zum klaren Verständnis nochmal festhalten, was die Wissenschaft unter Zwischenformen versteht. Wenn wir Lebewesen systematisch einordnen untersuchen wir ihre apomorphen Merkmale, die diese Lebewesen kennzeichnen und in ein hierarchisch verschachteltes System gemeinsamer Merkmale einordnen. Wenn sich neue Arten im Stammbaum von ihren Vorläuferarten abspalten, sind sie immer noch Teil dieser Verwandtschaft und damit immer noch Teil der höheren Ränge der Verwandtschaft. Die Art Mensch, Homo sapiens, gehört zur Ordnung der Primaten und alle Primaten zu den Säugetieren usw. Wenn neue Arten entstehen, spalten sie sich an ihrer untersten Stelle ab – der Bildung neuer Arten. Verfolgt man jedoch diese Artabspaltungen weiter in die Vergangenheit, zeigt sich, dass sie sich in immer größere Gruppen zusammenfassen lassen. Und: je weiter wir in die Vergangenheit diese Artabspaltungen zurückverfolgen, sehen wir, dass sich die frühen Vertreter einer Evolutionslinie immer ähnlicher sehen. Wir haben im Beitrag über die Kladisitik dies am Beispiel der Gruppe der Carnivora, also der Raubsäugetiere, wozu Hunde, Katzen und Bären gehören, dargestellt.
Kreationisten behaupten Hunde und Katzen können nur durch Evolution entstanden sein, wenn Hunde, von einer Generation zur nächsten, Katzenbabys auf die Welt bringen (oder andersrum) und eine Übergangsform zwischen beiden z. B. eine Katze mit Hundekopf (oder anders rum) wäre. Da aber Hunde immer nur Hunde zur Welt bringen und Katzen nur Katzen, gäbe es keine Evolution, wären sie fertige Grundtypen, die von Gott geschaffen wurden. Wir hingegen haben gezeigt, dass Hunde und Katzen über eine lange Kette von Verzweigungen, Seitenlinien und zigfachen Artabspaltungen im Verlauf von vielen Millionen Jahren auf einen gemeinsamen Vorfahren zurückgehen.
Alle 37 Arten, die zur Familie der Hunde (Canidae) gehören, wie Wolf, Rotfuchs, Waldhund und afrikanischer Wildhund, sowie alle 40 Arten der Familie der Katzen (Felidae), wie Tiger, Wildkatze und Puma, angehören, gehen auf einen gemeinsamen Vorfahren zurück, der weder ein Hund noch eine Katze war. Dieser Vorfahre verfügte über Merkmale, die beide Gruppen gemeinsam haben. Je weiter man diese Evolutionslinie verfolgt, desto eher gleichen sich die frühen Vertreter dieser beiden Stammlinien einander. Die Familie der Hunde geht auf fossile Arten wie Hesperocyon zurück, die Katzenartigen auf fossile Vertreter wie Pseudaelurus. Beide Arten – Hesperocyon und Pseudaelurus – sind für den Laien kaum zu unterscheiden. Untersucht man die Merkmale der beiden Vertreter, wird man aber zwei Dinge feststellen: zum einen haben sie viele Merkmale gemeinsam, zum anderen zeigen aber andere Merkmale – z. B. der Aufbau der Zähne, oder der Feinbau des Skeletts – dass Hesperocoyn schon Merkmale aufweist, die nur Hunden gemeinsam sind, während Pseudaelurus Merkmale aufweist, die typisch Katze sind.
Als sich beide Evolutionslinien abspalteten und die eine Linie in Richtung Hundeartige führte, die andere in Richtung Katzenartige, entwickelten sich unterschiedliche Merkmale weiter, die zu den heute lebenden Arten führten. Das ist das generelle Prinzip, wenn man Evolutionslinien zurückverfolgt. Verfolgt man die Evolutionsgeschichte von zwei beliebigen Arten zurück, wird man feststellen, dass ihre frühen Vertreter immer einander ähnlicher sehen, dass sie der Laie, teilweise noch nicht mal der Paläontologe, klar unterscheiden kann.
Leiter oder Busch?
Abb. 2: Evolution: Leiter oder Busch?
Unabhängig von der Unwissenheit der Kreationisten zum Thema Evolution und Übergangsformen gibt es jedoch ein weiteres populäres Missverständnis bezüglich der Evolutionsdarstellungen, die auch von jenen vertreten wird, die Kreationismus für reinsten Blödsinn halten.
Hier muss ich ein wenig ausführen:
Traditionell wird gerne von einer “Leiter” der Evolution gesprochen, bei der sich “primitive” Arten in “höher entwickelte” verwandeln. Man spricht auch gerne über die große Seinskette, die durch Bindeglieder miteinander verbunden wird. Sind diese Bindeglieder nicht bekannt, spricht man gerne vom „missing link“. Diese Bindeglieder stellen sogenannte Zwischenformen dar und Kreationisten lieben es auf die fehlenden Zwischenformen zu verweisen.
Die berühmteste Darstellung der Vorstellung der Evolution als Leiter ist die Entwicklungsreihe vom Affen zum Menschen. Als Übergänge sieht man dann Vertreter der Urmenschen wie Homo errectus und den Neandertaler (Abb. 3). Diese Darstellung erweckt den Anschein, als ob es eine Evolutionsreihe gab, die ohne weitere Abspaltungen vom primitiven Affen hin zum modernen Menschen führte. Fehlte ein solches Bindeglied vom Affen – oder allgemeiner vom Tier – zum Menschen, sprach man vom missing link.

Abb. 3: Menschliche Evolution als Leiter?
Die Bedeutung des missing links sollte also besagen, dass in der darwinschen Theorie eine entscheidende Verbindung zwischen Menschen und anderen Tieren, allen voran den Affen, fehlte. Das sagen die Kreationisten ja auch gerne: „Ihr habt noch nicht das fehlende Bindeglied zwischen Art X und Z gefunden“. Findet man doch eines, z. B. die Art Y sagen sie dann: „Aber nun habt ihr nicht das fehlende Bindeglied zwischen Art X und Y und das Bindeglied von Art Y und Z gefunden!“ Das würde immer so weiter gehen und bei jedem Bindeglied das gezeigt würde, entstünden immer zwei neue Lücken.
Doch das Problem des missing links ist ein anderes: Es ist der Mythos der großen Seinskette, wonach alle Dinge im Universum auf einer Leiter stehen. Ganz oben befindet sich Gott, dann die Engel, dann die Menschen, dann die Tiere, dann die Pflanzen und ganz unten schließlich die Steine und weitere unbelebte Dinge. Und natürlich stehen in dieser großen Seinskette nicht alle Menschen auf einer Stufe, denn Männer stehen angeblich höher als Frauen und die Weißen über den Schwarzen.
Dieser Mythos der großen Seinskette fand auch Einklang in die Vorstellungen zur Evolution im viktorianischen Zeitalter und hält sich bis heute hartnäckig in den Vorstellungen unserer Kultur. So hätten sich „niedere“ Tiere, sagen wir mal der Regenwurm, zu „höheren“ Tieren, sagen wir z. B. den Affen entwickelt. Und aus den Affen entwickelte sich der höher entwickelte Mensch. Das Problem dieser Vorstellung ist, dass sie eine zutiefst falsche und unevolutionäre ist. Denn mit welchem Recht können wir aus evolutionsbiologischer Sicht behaupten der Wurm ist primitiver als der Affe und dieser wiederrum als der Mensch? Hier geht es nicht darum, den Menschen „nur“ als irgendein Vieh abzustempeln, der über keine besonderen Merkmale verfügt, die ihn von allen anderen Lebewesen unterscheiden. Es geht um rein evolutionäre Fragestellungen: Der Erfolg der Evolution misst sich danach sich an die Umwelt anzupassen und seine Gene an die Nachkommen weiterzugeben. Weil heute Regenwurm, Schimpanse und Mensch existieren, haben sie sich auf ihre Weise unterschiedlich an die Umwelt angepasst und sich fortgepflanzt. Man könnte zwar einwenden, dass der Affe klüger ist als der Regenwurm, gleichzeitig kann man aber mit Fug und Recht behaupten, dass Regenwürmer in ihrer Fortpflanzung und Anpassung an die Umwelt viel erfolgreicher sind. Welche Maßstäbe setzt man also an, etwas aus Sicht der biologischen Evolution für höher oder niedriger entwickelt zu sehen? Weil alle heute lebenden Arten bis in unsere Gegenwart existieren, kann man daher davon ausgehen, dass sie ihre eigenen evolutionären Strategien hatten, die sie erfolgreich meistern konnten.
Es gibt also keine höheren oder niedrigen entwickelten Arten. Da wir alle auf einen gemeinsamen Vorfahren zurückgehen, haben alle heutigen Organismen, von der Bakterie hin zu uns Menschen, die gleiche Evolutionsgeschichte hinter uns. Man hat sich nur in unterschiedliche Richtungen hin entwickelt und spezialisiert. Somit gibt es auch keine “primitiven” und “höher” entwickelten Formen, es gibt nur ursprüngliche und abgeleitete Merkmale.
Wenn wir uns nochmal die Evolutionsleiter vom Affen zum Menschen anschauen, so ist sie ebenfalls aus einem anderen Aspekt falsch. Evolutionslinien verlaufen nicht in einer Linie, sondern sie verzweigen sich an vielen Stellen, sodass mehrere Entwicklungslinien parallel verlaufen. Gerade auch in Bezug zur Evolution des Menschen gab es viele Vormenschenarten, die gleichzeitig lebten. Das ist Folge des Evolutionsprozesses: einzelne Populationen spalten sich, z. B. in Form von geographischer Isolation, von der Gründerpopulation ab. So entstehen aus einer Population zwei oder mehr. Mit ausreichend Zeit und Isolation, also dass sich die Populationen nicht mehr vermischen können, entwickeln sie sich unabhängig voneinander weiter zu neuen Arten. Verschwinden die Isolationsbarrieren, können die Population wieder in Kontakt treten, vermischen sich aber nicht, es bilden sich also neue Arten. Wiederum können sich aus diesen Populationen durch Isolation weitere abspalten und neue Arten bilden, andere wiederrum sterben aus etc. durch diese Artabspaltung verläuft die Evolution also nicht auf einer Linie, oder Leiter, sondern als Busch mit vielen Abzweigungen, Seitenlinien, Parallelentwicklungen und Aussterbeereignissen.
Evolution ist also keine Leiter und kein linearer Prozess von niederen zu höher entwickelten Lebewesen. Die Evolution gleicht vielmehr einem Busch, der derselben Wurzeln entspringt, dessen Zweige sich aber in sämtliche Richtungen strecken. Er treibt ständig neue Äste hervor, alte sterben ab. Doch kein Zweig bevorzugt eine bestimmte Richtung. Arten können sich aufspalten und dann in verschiedene Richtungen weiterentwickeln. Insbesondere kann eine Entwicklungslinie neue Merkmale entwickeln (die heißen dann “Apomorphien”), während eine andere die basalen Plesiomorphien behält. Diese Erkenntnis wird wichtig, wenn wir über Übergangsformen sprechen.
Stellen wir aber erstmal beispielhaft ein typische Zwischenform dar: den Archaeopteryx.
Archaeopteryx als Übergangsform

Abb. 4: Archaeopteryx als Bindeglied
Der Archaeopteryx ist wohl das bekannteste sog. missing link, welches den Übergang von Dinosauriern zu Vögeln darstellt. Und es ist die Übergangsform, über die Kreationisten sehr gerne sprechen – und zwar sehr viel dummes Zeug.
Archaeopteryx ist eine Art Mosaikform aus abgeleiteten und ursprünglichen Merkmalen. Kreationisten sagen Archaeopteryx ist nur ein richtiger Vogel, weil er Federn und Flügel hat wie jeder Vogel. Der Urvogel besaß zwar „hundertprozentig echte“ Vogelmerkmale, wie etwa ein Federkleid, ein zum Gabelbein verwachsenes Schlüsselbein und ein nach hinten gerichtetes Schambein. Gleichzeitig besaß er aber noch nicht alle Merkmale der heute lebenden Vögel (wie etwa den Hornschnabel, verwachsene Finger usw.). Darüber hinaus kamen ihm, wie den heutigen Vögeln, typische „Reptilienmerkmale“ zu, aber auch solche, die den heutigen Vögeln fehlen (beispielsweise Bauchrippen, Kiefer mit Zähnen, einen langen Schwanz usw.)
Dieses Merkmalsmosaik weist dem Urvogel im phylogenetischen System ohne jeden Zweifel eine Position zu, die eine Zwischenstellung hinsichtlich der Hierarchie der heute lebenden Vögeln und den übrigen Sauropsiden („Reptilien“) einnimmt. Die Verwandtschaft und Merkmalsverteilung bei Vögeln und Raubdinosauriern sind so gut dokumentiert, dass man davon ausgehen muss, dass Vögel eine Gruppe der Dinosaurier sind. Genauer gesagt zeigt die Merkmalsverteilung, dass Vögel mehr Gemeinsamkeiten mit der Gruppe der fleischfressenden theropoden Dinosaurier haben, als diese theropode Dinosaurier mit anderen Dinosauriern. Dennoch bestehen Kreationisten darauf, dass Archaeopteryx zu 100% ein Vogel und keine Übergangsform sei. Doch jeder 100%ige Vogel hat einen Schnabel, Archaeopteryx nicht. Jedem 100%igen Vogel fehlen Zähne, Archaeopteryx hat welche. Jedem 100%igem Vogel fehlt eine Schwanzwirbelsäule, Archaeopteryx hat eine. Damit zeigen Kreationisten ihre Unehrlichkeit und Unwissenheit in der Argumentation. Aber damit nicht genug.
Andere Kreationisten, u. a. eine Amateurgruppe um den Astronomen Fred Hoyle, veröffentlichten eine Reihe von Artikeln, indem sie behaupteten, dass das Londoner und Berliner Exemplar des Archaeopteryx eine Fälschung seien. Sie unterstellten, dass Archaeopteryx kein Vogel, sondern ein gewöhnlicher Dinosaurier sei, bei dem die Federabdrücke im Fossil nachträglich aufgebracht wurden. d. h. anders als andere Kreationisten sagen sie: Archaeopteryx war nicht zu 100% Vogel, sondern zu 100% Dinosaurier, dem man mit falschen Federn geschmückt hat. Ich liebe es, wenn sich die Wissenschaftsgegner selbst nicht einig sind. Die Fälschung glaubt Hoyle übrigens nicht anhand der Untersuchung des aktuellen Fossils herausgefunden zu haben, sondern, indem er sich ein Foto des Londoner Exemplar anschaute. Seine Fälschungs-Hypothese stützt Hoyle vor allem auf „zwei Ungereimtheiten“, die er beim Betrachten der Fossil-Fotos entdeckte: Das feinkörnige Substrat, auf dem das Gefieder des Urvogels deutlich zu erkennen ist, habe eine ganz andere Konsistenz als der Rest des Abdrucks und sei wahrscheinlich mit einer Paste aus zermalenem Kalkstein auf ein echtes Reptilien-Fossil aufgetragen und dann die Federabdrücke hineinmanipuliert worden. Das Londoner Exemplar des Archaeopteryx besteht aus zwei Platten, die jeweils eine Skeletthälfte enthalten. Die Federabdrücke beider Platten seien von unterschiedlicher Tiefe und überdies nicht exakt deckungsgleich, so Hoyle.
Diese Unterstellungen Hoyles und anderer wurden jedoch von einer Forschergruppe des Natural History Museums of London widerlegt (Charig et al. 1986). Sie untersuchten dabei nicht das Foto, sondern arbeiteten mit dem Original-Fossil. Die angeblichen Beweise für die Fälschung konnten leicht widerlegt werden. Unregelmäßigkeiten in der Tiefe von Fossilabdrücken sind nichts Ungewöhnliches. Schließlich werden Fossilien nicht in einer modernen Industriepresse hergestellt, sondern waren über Jahrmillionen hinweg den unterschiedlichsten Drücken ausgesetzt. Mikroskopische Untersuchungen belegten zudem, dass die feinen Haarrisse in den Federn der zwei Teilstücke der Platte zueinander passen, genauso auch der Verlauf von Mangankristallen im Gestein. Eine viel spätere Studie machte vom Londoner Exemplar Aufnahmen mit einem Laserscanner. Die virtuellen Teilstücke des Archaeopteryx passen und fügen sich nahtlos in einer Skala zusammen, die in Mikrometern (Tausendstel Millimeter) gemessen werden kann.
Nicht zu vergessen: Als Hoyle seinen Artikel schrieb, gab es 5 Archaeopteryx-Exemplare, die zwar alle aus den Solnhofener Plattenkalken (oberer Jura) des mittleren Altmühltales in Bayern stammen, aber zu unterschiedlichen Zeiten, im Verlauf eines Jahrhunderts gefunden wurden – da müsste sich also ein langlebiger Witzbold über 100 Jahre hinweg mit dem Fälschen von Archaeopteryx-Fossilien beschäftigt haben. Mittlerweile sind 12 Exemplare gefunden worden.
Ende der Geschichte: Archaeopteryx ist echt, er ist eine Mosaikform, die den Übergang von Raubdinosauriern zu modernen Vögeln darstellt, Kreationisten haben keine Ahnung von Fossilien und sind sich noch nicht mal einig ob Archaeopteryx Dinosaurier oder Vogel war.
Weitere Uneinigkeiten
Diese Uneinigkeit seitens der Kreationisten findet sich übrigens nicht nur beim Archaeopteryx, sondern auch bei anderen Fossilfunden. So z. B. bei den Fossilfunden unserer eigenen Ahnenreihe. Für Kreationisten ist man nur entweder zu 100% Affe oder zu 100% Mensch, nichts dazwischen, den sogenannten Affenmenschen gebe es nicht. Diese Aussage ist schon unklug genug, denn wer Ahnung von Systematik hat, der weiß, dass wir Menschen zur Ordnung der Primaten gehören und damit Affen sind, etwas was schon der Entdecker der zoologischen Nomenklatur, Carl von Linne festgestellt hatte, obwohl er an die Schöpfung glaubte und Arten für nicht veränderbar hielt. Die Forderung nach einem Affenmenschen ist in etwa so sinnvoll, wie die Frage nach einem halb-Hund-halb-Dackel oder einem Halb-Mensch-halb-Wirbeltier.
An dieser Stelle soll die hoch interessante Fossilgeschichte der Hominiden nicht erörtert werden. Hier geht es vielmehr darum, wie Kreationisten nicht in der Lage sind, diese richtig zuzuordnen.
Drei „professionellen“ Kreationisten wurde unabhängig voneinander der Schädel eines Fossils gezeigt, der die Nummer KNM-ER 1470 trägt. Schauen Sie sich dieses Fossil genau an: Was sagen sie? Ist das zu 100% ein Affe (besser ein Affe, der kein Mensch ist) oder zu 100% ein Mensch?

Abb. 5: Der Schädel KNM-ER 1470 Affe oder Mensch?
Die Lösung:
Es handelt sich hierbei um den Schädel des Homo rudolfensis, eines sehr frühen Vertreters der Gattung Homo und lebte vor etwa 1,9 Mio. Jahren in Afrika. Wissenschaftler debattieren, ob es sich bei Homo rudolfensis nicht doch um einen Vertreter der Gattung Australopithecus gehört. Bei seiner Entdeckung im Jahr 1972 hatte man ihn auch als Vertreter der Art Homo habilis oder Kenyanthropus rudolfensis diskutiert. Wir haben es hier also mit einem Vertreter zu tun, der aus der Zeit stammte, als sich die Gattung der Australopithecinen weiter differenzierte und sich aus einigen dieser Gruppe die Gattung Homo entwickelte. Homo rudolfensis verfügt über Merkmale die typisch für beide Gattungen sind und das macht seine Einordnung so interessant. Wie haben die drei prominenten [Teletubbies … eh] Kreationisten getippt?
1979 wurde Duane Gish, dem damaligen Leiter des Institute for Creation Research, dieses Fossil gezeigt (ohne den Namen der Art zu erwähnen). Er identifizierte ihn als 100% menschlich. 6 Jahre später wurde ihm dasselbe Fossil nochmal gezeigt und diesmal hat er ihn als 100% Affe identifiziert. 1992 hatte ein anderer Kreationist, Marvin Lubenow dieses Fossil als 100% menschlich angesehen, während sein Kollege A. W. Mehlert es 1996 als 100% Affe ansah (Aron Ra 2016). Braucht es da noch mehr Beweise für die absolute Inkompetenz der Kreationisten?
Die Suche nach den Vorfahren
Manche Kreationisten kommen dann jedoch mit einem anderen „Argument“: Sie sagen: ja mag sein, dass Archaeopteryx Merkmale von Dinosauriern und Vögeln gemeinsam hat, aber er ist nicht der Vorfahre aller Vögel, aufgrund diverser Details in der Anatomie.
Dies ist ein netter Taschenspielertrick, der aber bei genauerer Betrachtung nicht im Widerspruch stehen muss. Findet man Mosaik- und Übergangsformen heißt nicht automatisch, dass diese die direkten Vorfahren sind, sondern vielleicht Seitenäste in der Entwicklung. Die Erklärung dafür ist ganz einfach.
Über 99% aller Lebewesen sind im Verlauf der Erdgeschichte ausgestorben und nur ein winziger Bruchteil davon wird zum Fossil. Um mal mit Zahlen zu bombadieren: heute sind etwa 2 Mio. Tier- und Pflanzenarten beschrieben worden. Die tatsächliche Zahl dürfte aber fast astronomisch größer sein, denn regelmäßig werden neue Arten entdeckt und beschrieben. Schätzungen der Arten die heute unsere Erde bevölkern liegen zwischen 3 Mio. und 100 Mio., vielleicht sogar noch mehr. Wenn wir davon ausgehen, dass 10 Mio. Arten unsere heutige Erde bevölkern und im Verlauf der Erdgeschichte 99% aller Arten ausgestorben sind, brachte die Evolution 1 Milliarde unterschiedlicher Arten hervor! Mindestens! Doch wie viele dieser ausgestorbenen Arten kennen wir als Fossilien? Bis 1999 sind rund 250.000 fossile Arten wissenschaftlich beschrieben worden (Prothero 1999), lassen wir es, da wir nun 2020 haben 300.000 bekannte fossile Arten haben. Das heißt nur ein Bruchteil aller Arten wird überhaupt zum Fossil und es wird nicht jedes Fossil entdeckt. Folgerichtig, wenn nicht alle Arten fossilisieren und wir nicht alle Fossilien entdecken, ist es sehr vage zu behaupten Fossil X ist der Vorfahre von der heute lebenden Art Y. Vielleicht kann es der Vorfahre sein, vielleicht aber nur ein entfernter Verwandter. Wir können hierzu auch unseren eigenen Familienstammbaum als Vergleich nehmen. Wir sind das Produkt unserer Eltern. Aber unsere Eltern haben meist Geschwister (unsere Tanten und Onkel), die ebenso Kinder haben. Wiederum kommen noch die Großeltern hinzu, die ebenfalls Geschwister haben usw. usf. Wenn jemand (z. B. ein Ahnenforscher) meinen Familienstammbaum untersuchen will und meine Cousine, meine Großtante oder meine Urgroßmutter ausfindig machen kann, nicht aber meine Eltern, so ist es offensichtlich, dass diese nicht meine Eltern sein können. Dennoch ist nicht abzustreiten, dass sie mit mir verwandt sind. Und nur weil man meine Eltern nicht findet, heißt es nicht automatisch, dass ich durch einen göttlichen Akt geschaffen wurde. Genauso verhält es sich mit dem Fossilfund. Wenn wir Fossilien finden und sie verwandtschaftlich in die Nähe rezenter Arten rücken, sagen wir damit nicht, dass wir genau den Vorfahren entdeckt haben, der direkt zur untersuchten heute lebenden Art führte. Wir sagen lediglich, dass dieses Fossil eine Reihe von Merkmalen mit heute lebenden Arten hat, die auf eine gemeinsame Abstammung hindeuten. Es ist, wenn überhaupt, dann ein “Vorfahre” im weitesten Sinne, sodass wir z. B. sagen können, Vögel entstammten einer Gruppe von Raubdinosauriern und Archaeopteryx ist dafür ein Beispiel. Wir sagen aber nicht, die Art Archaeopteryx lithographica ist der Vorfahre aller heute lebenden Vögel.
Bei der Dokumentation der evolutionären Übergänge geht es nicht primär um Genealogie, also nicht darum, welche Art sich zu welcher entwickelt hat, sondern welche evolutionären Merkmalsveränderungen stattfinden müssen, um diese zu erklären. Hierzu kann man auch einen entfernteren verwandten zu Rate ziehen. Wie bei unserem Stammbaum: auch durch die Befragung unserer Onkel und Tanten können wir etwas über unsere eigene Familiengeschichte erfahren, wir müssen also nicht immer die Eltern finden.
Wenn Evolutionsbiologen also von Vorfahren oder Zwischenformen sprechen, meinen sie, dass sich aus einer Gruppe von Arten eine Gruppe von anderen Arten entwickelt hat. Dies geschieht anhand der Analyse plesiomorpher und apomorpher Merkmale, also Anhand von Homologien.
Gruppen von Übergängen
Tatsächlich unterscheiden Paläontologen verschiedene Formen der Übergänge zwischen den Gruppen der Lebewesen.
Kathleen Hunt hat auf der Seite talkorigins.org zwischen 1994 und 1997 eine Liste fossiler Übergangsformen bei Wirbeltieren erstellt. Der Titel dieser Liste trägt den Namen „Transitional Vertebrate Fossils FAQ“. Bedenkt, dass diese Seite mittlerweile über 20 Jahre alt ist und sich in diesen zwei Jahrzehnten viel in der Paläontologie getan hat, gerade auch was Neuentdeckungen bei Fossilien angeht. Dennoch ist diese Liste eine schöne Quelle, für diejenigen, die Interesse daran haben. In ihrer Liste stellt sie aber drei unterschiedliche Typen der Übergänge bei Wirbeltieren dar:
-
Allgemeine Abstammungslinien
Dies ist eine Sequenz ähnlicher Gattungen oder Familien, die eine ältere Gruppe mit einer ganz anderen jüngeren Gruppe verbindet. Jeder Schritt in der Sequenz besteht aus einigen Fossilien, die eine bestimmte Gattung oder Familie darstellen, und die gesamte Sequenz erstreckt sich häufig über einen Zeitraum von mehreren zehn Millionen Jahren. Eine solche Linie zeigt offensichtliche morphologische Zwischenprodukte für jede größere strukturelle Veränderung, und die Fossilien kommen ungefähr (aber oft nicht genau) in der erwarteten Reihenfolge vor. Normalerweise gibt es immer noch Lücken zwischen den einzelnen Gruppen – nur wenige oder keine der Artbildungsereignisse bleiben erhalten. Manchmal wird nicht angenommen, dass die einzelnen Exemplare direkt von den nächstjüngsten Fossilien abstammen (wegen der oben genannten Gründe der Seltenheit, dass einzelne Arten zu Fossilien werden). Es wird jedoch angenommen, dass sie eng mit dem tatsächlichen Vorfahren verwandt sind, da sie im Vergleich zu den nächstältesten und nächstjüngsten „Verbindungen“ eine mittlere Morphologie aufweisen. Der Hauptpunkt dieser allgemeinen Abstammungslinien besteht darin, dass Tiere mit mittlerer Morphologie zu geeigneten Zeiten existierten und somit die Übergänge von den vorgeschlagenen Vorfahren vollständig plausibel sind. Allgemeine Abstammungslinien sind für fast alle modernen Gruppen von Wirbeltieren bekannt. So die Evolution der Raubdinosaurier zu modernen Vögeln oder die frühen Synapsiden zu den modernen Säugetieren.
-
Übergänge von Art zu Art:
Dies ist eine Reihe von zahlreichen einzelnen Fossilien, die einen Wandel von einer Art zu einer anderen zeigen. Es ist eine sehr feinkörnige Sequenz, die das eigentliche Artbildungsereignis dokumentiert und normalerweise weniger als eine Million Jahre umfasst. Diese Übergänge von Art zu Art sind unverkennbar, wenn sie gefunden werden. Manchmal treten diese Sequenzen nur in einem begrenzten geografischen Gebiet auf (dem Ort, an dem die Artbildung tatsächlich stattgefunden hat), wobei Analysen aus jedem anderen Gebiet eine scheinbar „plötzliche“ Änderung zeigen. In anderen Fällen kann der Übergang jedoch über ein sehr weites geologisches Gebiet beobachtet werden. Viele „Art-zu-Art-Übergänge“ sind bekannt, hauptsächlich für wirbellose Meerestiere und jüngere Säugetiere (beide Gruppen haben tendenziell gute Fossilienbestände), obwohl sie nicht so häufig vorkommen wie die allgemeinen Abstammungslinien.
Hier sollen nur einige Beispiele für solche Art-zu-Art-Übergänge genannt werden.
Eine Arbeit, veröffentlicht in einer Symposiums-Schrift des französischen Paläontologen Jean Chaline beschreibt die graduelle Veränderung der frühen Vertreter der Katzenartigen von Haplogale media über Proailurus lemansis, zu Pseudaelurus transitorius zu Ps. lorteti zu Ps. rmoieviensis zu Ps. quadridentatus. Diese europäischen Linien waren die Vorfahren jener Linien die zu den Luchsen und Großkatzen führten. Andere Entwicklungen fanden in Nordamerika, Afrika und Asien statt (Chaline 1983).
Der Wirbeltierpaläontologe Björn Kurten schrieb 1976 ein Buch über die Evolutionsgeschichte der eiszeitlichen Höhlenbären. Dort stellt er die Evolution von frühen Formen wie Ursus minimus, der vor 5 Mio. Jahren lebte, bis hin zum Höhlenbären (Ursus spelaeus) dar. Dieser Übergang ist gekennzeichnet durch eine nahezu lückenlose Fossilienreihe mit Artübergängen wie Ursus etruscus und Ursus savini. Dabei ist dieser Übergang so graduell, dass es schwierig ist, zwischen den einzelnen Arten eine Grenze zu ziehen. Die frühe Form Ursus minimus gilt dabei auch als der Vorfahre der heute lebenden Bärenarten wie Braun- und Eisbär (Kurten 1976).
Es sei noch ein drittes Beispiel genannt. Vincent Maglio veröffentlichte 1973 eine Arbeit zur Evolutionsgeschichte der Familie der Elefanten (Familie Elephantidae). Er konnte zeigen, dass mindestens 7 der damals 17 bekannten Arten der Elefantenfamilie aus dem Quartär (jene Zeitepoche, die vor 2,6 Mio. Jahren begann) durch eine reibungslose Ahnenreihe aus ihren Vorfahren entstanden sind. Er zeigte auf, dass die Art Elephas recki durch eine fortschreitende Reihe von Entwicklungsstadien zurückverfolgt werden kann, die fast unmerklich ineinander übergehen und aus denen die Art Elephas iolensis entstand. Auch wenn beide Arten voneinander zu unterscheiden sind, zeigen sich im Fossilbestand viele Übergänge, bei der Elephas iolensis als ein weit fortgeschrittenes Evolutionsstadium von Elephas recki angesehen werden kann. Maglio dokumentierte auch reibungslose Übergänge zwischen drei europäischen Mammutarten: Mammuthus meridionalis –> M. armeniacus –> M. primigenius (Maglio 1973, Abb. 6). Diese Entwicklungsreihe der europäischen Mammuts wurde von Adrian Lister 1993 bestätigt und diese graduelle Evolution der Mammuts konnte er auch für die nordamerikanischen Arten bestätigen (Lister 1993).
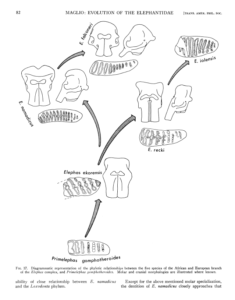
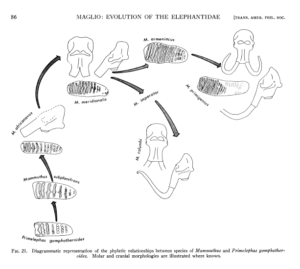
Abb. 6: Evolution der Elefanten nach Maglio (1973)
Wir sehen also: bei einigen Fossilien zeigen sich sogar Übergänge von einer Art zur nächsten. Ich habe mich dabei auf große Wirbeltiere konzentriert, weil es die meisten eher interessiert als das Kleinvieh. Tatsächlich zeigen sich aber die besten Art- zu Artübergänge bei sogenannten Mikrofossilien. Hierzu zählen z. B. die Foraminiferen (Abb. 7). Das sind eukaryotische Einzeller, sie haben also einen Zellkern, die hauptsächlich im Meer leben und ein hartschaliges Gehäuse, meistens aus Kalk bestehend, bilden. Die verschiedenen Arten bilden die unterschiedlichsten Gehäuse aus und diese sind ein gutes diagnostisches Merkmal zur Unterscheidung der Arten. Und nun kommts: heute leben etwa 10.000 Foraminiferenarten, die Zahl der fossilen Arten beträgt etwa 40.000, ist also viermal so hoch. Und es sind mit die häufigsten Fossilien, die man findet. Und bei der Menge und Häufigkeit der Foraminiferen-Fossilien lassen sich hier Art- zu Artübergänge besonders gut dokumentieren. Nebenbei: der Löwenanteil der Paläontologen arbeitet mit Mikrofossilien und nicht mit Mammuts, Urmenschen und Dinosauriern.

Abb. 7: Foraminiferen
3. Übergänge zu neuen höheren Taxa
Wie in der FAQ von Kathleen Hunt zu sehen ist, führen beide Arten von Übergängen häufig zu einem neuen „höheren Taxon“ (eine neue Gattung, Familie, Ordnung usw.). Daran ist nichts Magisches. Die ersten Mitglieder der neuen Gruppe sind keine bizarren, chimären Tiere; sie sind einfach eine neue, etwas andere Art, die sich kaum von den Elternarten unterscheidet. Schließlich führen sie zu einer anderen Art, was wiederum zu einer weiteren Art führt, und so weiter, bis sich die Nachkommen radikal vom ursprünglichen Elternbestand unterscheiden. Wir haben das zu Beginn am Beispiel der Hunde und Katzen erläutert. Ein anderes Beispiel stellen Huftiere dar. Zum Beispiel können die Ordnungen Perissodactyla (Unpaarhufer wie Pferde, Nashörner und Tapire usw.), die Ordnung der Artiodactlya (Paarhufer wie Rinder, Antilopen und Hirsche) und die Ordnung Cetacea (Wale) auf frühe eozäne Tiere zurückgeführt werden, die sich nur geringfügig voneinander unterschieden und überhaupt nicht wie Pferde, Kühe oder Wale aussahen. (Sie sahen eher aus wie kleine Füchse mit waschbärenähnlichen Füßen und einfachen Zähnen.) Aber in den folgenden zehn Millionen Jahren wurden die Nachkommen dieser Tiere immer unterschiedlicher, und jetzt teilen wir sie in drei verschiedene Ordnungen ein, obwohl ihre Stammformen kaum voneinander zu unterscheiden sind.
Dies sind nur einige Beispiele, die zeigen, dass auch die Übergänge von einer Gruppe zur nächsten im Fossilbestand gefunden werden kann. Kathleen Hunts Liste zeigt noch wesentlich mehr Beispiele. Wenn man die Fossilien kennt, wenn man die Arbeit der Paläontologen studiert, wird man feststellen, dass der Fossilbestand eine ganze Reihe an Übergangsformen von einer Gruppe zur nächsten aufzeigt. Kreationisten verschweigen diese Tatsache, da die einzigen Fossilien, die sie kennen, Spielzeugdinosaurier aus dem Supermarkt sind.
One of these things is not like the other
Diese Unwissenheit über Fossilien seitens der Kreationisten kann aber noch stärker ins Absurde geführt werden. Eine Lieblingsmethode von Ihnen ist es zu zeigen, dass sich die fossilen Arten nicht geändert haben. Dabei zeigen sie ein Bild eines Fossils und daneben eines heute lebenden Tieres und sagen: schaut euch die beiden an, sie haben sich überhaupt nicht verändert. Besonders skuril wird es aber dann, wenn Fossilien mit heute lebenden Tieren verglichen werden, die gar nichts miteinander zu tun haben. So u. a. im Buch Atlas of Creation des islamischen Kreationisten Harun Yahya. Dieses extrem schwere Hochglanzbroschüre, mit dem man ein ordentliches Workout machen kann, wurde von Yahya kostenlos an Universitäten und Schulen verschickt. Richard Dawkins hat sich das Buch Atlas of Creation vorgenommen und in einem Vortrag auf humorvolle Weise bloßgestellt.
Harun Yahya ist bei weitem nicht der einzige. Auf Facebook entdeckte ich eine Seite, die dasselbe macht. Titel der facebookseite ist „Where is the evolution“. Auch hier werden Fossilien neben modernen Tieren gezeigt, um zu demonstrieren, dass keine Evolution stattgefunden hat. Die Behauptungen auf dieser Seite sind so dämlich konzipiert, dass ich mich frage, ob es sich um eine Satire-Seite handelt. Da aber Kreationisten die dümmlichsten Aussagen für wahr halten, ist es durchaus denkbar, dass dies ernst gemeint ist. Die Krone der Absurdität zeigt dieses Bild:

Links sehen wir das Fossil eines Psittacosaurus, rechts das Skelett eines modernen Papageis. Das Bild behauptet, beide seien gleich, Evolution fand nicht statt. Ich muss mir hier ernsthaft die Frage stellen, was der Ersteller dieses Bildes geraucht hat. Wenn man bedenkt, Harun Yahyas Angelhaken können nicht mehr übertroffen werden, so werde ich hier eines Besseren belehrt. Psittcaosaurus heißt auf Deutsch Papageiechse. Er gehört zu den Dinosauriern und ist ein früher Vertreter jener Gruppen, zu denen z. B. der Triceratops gehört. Lebte aber nicht vor 227 Mio. Jahren, sondern vor 126 Mio. Jahren. Der Name kommt daher, dass dieser Dinosaurier einen Schnabel hatte, der dem von Papageien ähnlichsieht. Aber keiner, wirklich keiner hat jemals behauptet, Papageien stammen vom Psittacosaurus ab. Selbst innerhalb der Gruppe der Dinosaurier gehören sie jener an, die sich nicht zu Vögeln entwickelt haben! Ein wenig googlen reicht schon, um zu wissen, dass beide nicht dasselbe sein können.
Was können wir zusammenfassen: Evolution ist eine Tatsache, die auch im Fossilbericht gut dokumentiert ist. Kreationisten hingegen habe null Plan von Evolution und was evolutionäre Übergänge angeht – und sie sind sogar zu blöd die einfachsten Grundlagen der Zoologie zu verstehen, sonst würden sie Angelhaken nicht mit Fliegen vergleichen.
Literatur:
Aron Ra (2016): Foundational Falsehoods of Creationism. Pitchstone Publishing
Chaline, J. 1983. Modalites, Rythmes, Mecanismes de L’Evolution Biologique: Gradualisme phyletique ou equilibres ponctues? Editions du Centre National de la Recherche Scientifique, Paris. [collection of symposium papers, most in French with English abstracts provided, some in English.]
Charig AJ, Greenaway F, Milner AC, Walker CA, Whybrow PJ (1986): Archaeopteryx is not a forgery. Science. 1986;232(4750):622‐626. doi:10.1126/science.232.4750.622 https://pubmed.ncbi.nlm.nih.gov/17781413/
Hoyle, F.; Wickramasinghe, N. C.; Watkins, R. S. (1985). „Archaeopteryx“. British Journal of Photography. 132: 693–694.
Hunt, K. (1997): Transitional Vertebrate Fossils FAQ http://www.talkorigins.org/faqs/faq-transitional.html
Kurten, B. 1976. The Cave Bear Story. Columbia University Press, New York.
Lister, Adrian. (1993). Evolution of mammoths and moose: The Holarctic perspective. Morphological Change in Quaternary Mammals of North America. 178-204.
Maglio, V.J. 1973. Origin and evolution of the Elephantidae. Trans. Am. Phil. Soc., New Ser. 63:1-149.
Miller, Keith B. n.d. Taxonomy, transitional forms, and the fossil record. http://www.asa3.org/ASA/resources/Miller.html
Nedin, C. (1999): All about Archaeopteryx http://www.talkorigins.org/faqs/archaeopteryx/info.html
Neukamm, M. (2001): Die Evolutionstheorie und der moderne Antievolutionismus http://www.martin-neukamm.de/junker.html
Patterson, Bob. 2002. Transitional fossil species and modes of speciation. http://www.origins.tv/darwin/transitionals.htm
Prothero, D. (1999): Fossil Record. In: Ronald Singer (Hrsg.): Encyclopedia of Paleontology. Fitzroy Dearborn Publishers, Chicago 1999, ISBN 1-884964-96-6, S. 491.
Prothero, D. (2017): Evolution – What the Fossils say and why it matters. Second edition. New York: Columbia University Press
TalkOrigina: Archaeopteryx http://www.talkorigins.org/indexcc/CC/CC214_1_1.html
Thompson, Tim. 1999. On creation science and transitional fossils. http://www.tim-thompson.com/trans-fossils.html
Watkins, R. S.; Hoyle, F.; Wickrmasinghe, N. C.; Watkins, J.; Rabilizirov, R.; Spetner, L. M. (1985). „Archaeopteryx – a photographic study“. British Journal of Photography. 132: 264–266.
Watkins, R. S.; Hoyle, F.; Wickrmasinghe, N. C.; Watkins, J.; Rabilizirov, R.; Spetner, L. M. (1985). „Archaeopteryx – a further comment“. British Journal of Photography. 132: 358–359, 367.
Watkins, R. S.; Hoyle, F.; Wickrmasinghe, N. C.; Watkins, J.; Rabilizirov, R.; Spetner, L. M. (1985). „Archaeopteryx – more evidence“. British Journal of Photography. 132: 468–470.
facebookseite zu „Where is the evolution“: https://www.facebook.com/ScienceExposingLies/
