
Das System von Linne hat einige große Schwächen:
Linné war kein Anhänger der Evolution, sondern glaubte an die Konstanz der Arten. Linné ordnete die verschiedenen Tiere und Pflanzen nach ihren Merkmalen, evolutionäre Ursprünge kamen ihm dabei nicht in den Sinn.
Dass Evolutionsverlauf und Verwandtschaftsbeziehungen früher keine große Rolle spielten, zeigt sich z. B. darin, dass Elefanten, Flusspferde und Nashörner als Dickhäuter (Pachydermes) zusammengefasst wurden, obwohl wir heute wissen, dass alle drei Gruppen unterschiedlichen evolutionären Linien entsprangen. Elefanten sind näher mit den Schliefern und Seekühen verwandt, Nashörner werden mit den Pferden und Tapiren zu den Unpaarhufern zusammengefasst und Flusspferde sind Paarhufer, deren nächsten heute lebenden Verwandten die Wale sind.
Es braucht also ein Ordnungsprinzip, dass den Evolutionsverlauf und die Verwandtschaftsbeziehungen berücksichtigt, denn nicht Merkmale definieren Gruppen, sondern die Abstammung. Es muss also untersucht werden, welche Merkmale geeignet sind die Verwandtschaftsbeziehungen zu ermitteln. Hier bedient man sich der Methoden der phylogenetischen Systematik.
In den 1960er Jahren hatte der Biologe Willi Hennig (1950, 1966) ein System ausgearbeitet, welches die verwandtschaftlichen Beziehungen zwischen den Lebewesen anhand von Merkmalsvergleichen ermitteln konnte. Man spricht von phylogenetischer Systematik oder Kladistik. Was ist das Besondere an dieser Methode im Vergleich zu anderen?
Hennigs wichtige Schlussfolgerung war, dass die Merkmale, die ein Lebewesen besitzt, nicht gleichwertig sind. Jedes Lebewesen ist ein Mosaik aus sogenannten fortgeschrittenen Merkmalen (auch Apomorphien genannt) oder ursprünglichen Merkmalen (auch Plesiomorphien genannt). Apomorphien sind Merkmale die nur die entsprechenden Lebewesen haben, Plesiomorphien sind jene, die man von den Vorfahren vererbt bekommen hat (vgl. Abb.1).

Abb. 1: Definition von Apomorphie und Plesiomoprhie nach Hennig
Dies kann am Beispiel der Klasse der Säugetiere verständlich gemacht werden (vgl. Abb. 2).



Abb. 2: Vielfalt der Säugetiere
Säugetiere müssen, um sie von anderen Gruppen zu unterscheiden, Merkmale aufweisen, die nur ihnen eigen sind, also ihre Apomorphien haben. Wir müssen also nach Merkmalen suchen, die Säugetiere als Gruppe gemeinsam haben und definieren; man sie also anhand dieser Merkmale von anderen Tiergruppen klar unterscheiden kann. Gemeinsame Merkmale einer Gruppe bezeichnet man auch als Autapomorphien.
Welche Merkmale eignen sich?
Säugetiere haben eine Wirbelsäule. Doch eine Wirbelsäule kommt auch bei anderen Tiergruppen vor, z. B. den Vögeln. Hierbei handelt es sich also um ein plesiomorphes Merkmal. Hingegen sind Merkmale wie Haare, Schweißdrüsen und die Produktion von Milch typisch für Säugetiere und zwar NUR für Säugetiere. Folgerichtig handelt es sich um apomorphe Merkmale, die die Säugetiere kennzeichnen.
Anhand solcher Merkmalsanalysen, bei denen die Autapomorphien der Lebewesen ermittelt werden, entstehen entsprechende Diagramme, die man als Kladogramm bezeichnet. Kladogramme sagen aus, wer mit wem näher verwandt ist (Abb. 3).
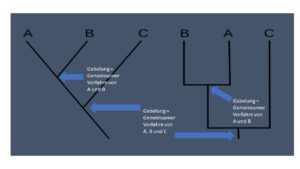
Abb. 3: Kladogramm
Bei einem Kladogramm zeigt sich ein für ihn typisches Verzweigungsmuster (Vgl. Abb. 3). Bei einer Verzweigung gibt es immer nur zwei Äste (man spricht von dichotomer Verzweigung). Jeder Ast ist durch ein abgeleitetes Merkmal begründet. Was dieses Merkmal jeweils sein soll, ist Gegenstand der Forschung. Der Verzweigungspunkt (Gabelung) zwischen den zwei Ästen stellt den gemeinsamen Vorfahren dar. Um es durch ein Beispiel am Kladogramm der Säugetiere zu verdeutlichen (Abb. 4):
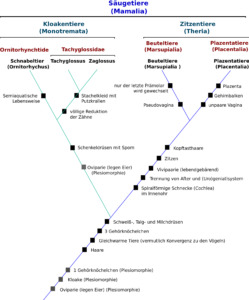
Abb. 4: Kladogramm der Säugetiere
Wie wir sehen, teilt man die Säugetiere in zwei Unterklassen ein: die Kloakentiere (Monotremata), wozu u. a. das Schnabeltier gehört und die Zitzentiere (Theria), welche Beuteltiere (Marsupialia) und Plazentatiere (Placentalia) miteinander vereinen.
Beginnen wir mit dem Stammbaum ganz unten, also an der Wurzel. Dort sind einige Merkmale aufgelistet, die der gemeinsame Vorfahre aller Säugetiere hatte: z. B. Haare, drei Gehörknöchelchen, sowie Schweiß- und Milchdrüsen. Danach spaltet sich das Kladogramm der Säugetiere in zwei Hauptäste, einer führt (der grüne Ast) zu den Kloakentieren, die sich wiederrum in zwei Äste aufspalten: Schnabeltiere und Schnabeligel. Die Schnabeligel teilen sich wieder in zwei Äste auf. Jede neue Abspaltung ist für nur diese Zweige typischen Apomorphien definiert. Der blaue Ast zeigt, dass Beuteltiere und Plazentatiere einen gemeinsamen Verzweigungspunkt haben, also auf einen gemeinsamen Vorfahren zurückgehen. Der gemeinsame Vorfahre verfügte über Merkmale, die Beuteltiere und Plazentatiere gemeinsam haben, so z. B. das Vorhandensein von Brustwarzen. Diese Apomorphie (neben anderen) definiert die Gruppe der Zitzentiere, zu denen Beutel- wie Plazentatiere gehören. Die gemeinsamen Merkmale von Beutel- und Plazentatieren hatten die Vorfahren dieser Gruppe, bevor sich diese in die zwei benannten Gruppen gespalten haben. Der gemeinsame Vorfahre spaltete sich dann aber ab in zwei Äste, die jeweils durch den Erwerb jeweiliger Apomorphien gekennzeichnet sind. So haben Beuteltiere eine sogenannte Pseudovagina und Plazentatiere eine Plazenta.
Natürlich werden in diesem Stammbaum nicht alle Apomorphien dargestellt und man sollte nicht vergessen, dass es 320 Beuteltierarten und etwa 6.000 Arten von Plazentatieren gibt. All diese Arten werden jeweils durch für sie typische Apomorphien gekennzeichnet, die sich in einem Kladogramm durch weitere Verzweigungen darstellen würden. Natürlich sollte auch nicht vergessen werden, dass wir uns nur einen kleinen Teil des Stammbaums des Lebens anschauen: hier werden die anderen Wirbeltiere, die Insekten, Würmer, Pflanzen, Pilze und Bakterien gar nicht berücksichtigt. Dieses Beispiel diente also nur der Vereinfachung.
Verzweigungen, die von einem gemeinsamen Verzweigungspunkt entstehen bezeichnet man auch als monophyletisch. Wenn wir uns das Kladogramm der Säugetiere nochmals vergegenwärtigen, bleibt die Frage:
Wo würden wir als Menschen einordnen? Wenn wir die Merkmale ansehen, so haben wir Haare, sind gleichwarm, haben Schweiß- und Milchdrüsen, sowie mit Hammer, Amboss und Steigbügel drei Gehörknöchelchen. Wir sind also Säugetiere.
Wenn wir die Merkmale weiterverfolgen, so haben wir eine Cochlea im Innenohr, unsere After- und Genitalöffnungen sind getrennt (wir haben also keine Kloake), wir sind lebendgebärend und haben Brustwarzen. Damit gehören wir auch zu den Zitzentieren (Theria). Auch haben wir, bzw. nur Frauen, eine Plazenta du eine unpaarige Vagina. Außerdem haben wir einen Gehirnbalken. Damit zählen wir auch zu den Plazentatieren. Es wird aufgefallen sein, dass in diesem Kladogramm als Apomorphien aller Zitzentiere auch die Kopftasthaare, also die Schnurrhaare, wie wir sie von Katzen und anderen Säugern kennen, genannt werden, die aber Menschen nicht haben. Im Verlauf der Evolution kann es vorkommen, dass ein gemeinsames Merkmal wieder verlorengeht. Dieses Nicht-Vorhandensein eines Merkmals, in unserem Fall das Fehlen von Schnurrhaaren, heißt nicht, dass wir dadurch keine Säugetiere sind, sondern sich weitere Spezialisierungen entwickelt haben, die durch die natürliche Selektion begünstigt worden sind. Somit ist das Rückbilden von Merkmalen, die ggfs. auch als rudimentäre Organe vorliegen können, ebenfalls Teil des Evolutionsgeschehens, weil diese zu neuen Anpassungen führen. Wale verfügen über keine Hintergliedmaßen und sind Wasserbewohner. Trotzdem handelt es sich bei ihnen nicht nur um Säugetiere, sondern auch um Tetrapoden, vierbeinigen Landwirbeltiere. Schlicht und einfach, weil sie einer Gruppe entstammten, die landlebende Säugetiere mit vier Beinen waren, die sich durch den Verlust ihrer Hintergliedmaßen auf das Leben im Wasser spezialisiert haben. Damit können wir eine weitere Regel der phylogenetischen Systematik ableiten: you can’t lose your ancestry – Man kann seine Verwandtschaft niemals loswerden.
Unsere jüngsten Vorfahren, die wir haben, sind unsere Eltern. Mit diesen teilen wir mehr Gemeinsamkeiten als mit unseren Cousins. Aber mit unseren Cousins haben wir mehr Gemeinsamkeiten, als mit unseren Nachbarn, Klassenkameraden usw., also Menschen, die nicht Teil unserer engen Verwandtschaft sind. Wir müssen uns aber darüber im Klaren sein, dass sie immer noch mit uns irgendwie verwandt sind – eben als Art Mensch.
Egal wie sehr wir uns von unseren Eltern und allen anderen Menschen unterscheiden mögen, sind wir immer noch Teil dieser Verwandtschaft, die wir nicht ablegen können.
Kreationisten reagieren oft mit der nicht begründbaren Ausrede, dass z. B. neu entwickelte Fliegenarten immer noch Fliegen sind, neue Fischarten immer noch Fische, neue Affenarten immer noch Affen usw. Na klar ist das der Fall! Aus Sicht der Evolution ist nichts anderes zu erwarten! Die Evolution konnte nicht zulassen, dass sie etwas anderes waren. Die biologische Klassifizierung ist hierarchisch. Wenn neue Arten entstehen, spalten sie sich an ihrer untersten Stelle ab – der Bildung neuer Arten. Also ist jede neue Art, die sich von einer ursprünglichen Fliegenart abspaltet, egal wie groß die Unterschiede sind, immer noch eine Fliegenart. Aber Fliege ist nicht gleich Fliege, so sind 150.000 verschiedene Fliegen- und Mückenarten beschrieben worden – eine sehr große Diversität. Würden wir aber die Abspaltungen weiter zurückverfolgen, tiefer in die Vergangenheit gehen, würden die Vorfahren unserer vorhin abgespaltenen neuen Fliegenart sich so stark unterscheiden, dass sie mit heute lebenden Fliegenarten weder kreuzbar wären, noch überhaupt für so etwas wie Fliegen gehalten werden würden.
Von Hunden und Katzen
Dasselbe gilt für das noch absurdere Vorurteil der Kreationisten, sie würden nur an Evolution glauben, wenn Hunde Katzenbabies zur Welt bringen. Auch das wird von der Evolutionstheorie nicht gefordert. Über eine lange Kette von Verzweigungen, Seitenlinien und zigfachen Artabspaltungen im Verlauf von vielen Millionen Jahren, können wir ermitteln, dass die heute lebenden Katzen- und Hundearten auf einen gemeinsamen Vorfahren zurückgehen. Dieser war nicht ansatzweise ein Hund oder eine Katze, sondern verfügte über Merkmale, die beide Gruppen gemeinsam hatten. Als sich beide Evolutionslinien abspalteten und die eine Linie in Richtung Hundeartige führte, die andere in Richtung Katzenartige, entwickelten sich unterschiedliche Merkmale, die zu den heute lebenden Arten führten. Zu Beginn dieser Verzweigung jedoch sahen sich die frühen Vertreter der beiden Evolutionslinien so ähnlich, dass se kaum zu unterscheiden waren. Die frühesten und urtümlichsten Vertreten der Carnivora (der Ordnung zu denen Hunde, Katzen, Marder und Bären gehören) waren die Miaciden (Abb. 5), die ihrer äußeren Erscheinung nach einem Wiesel ähnelten, bei genauerer Betrachtung jedoch Merkmale aufweisen, die alle Carnivora gemeinsam haben. 
Abb. 5: Vertreter der Gruppe der Miaciden, den frühesten Vertretern aus der Ordnung der Carnivora
Aus solchen Stammformen entwickelten sich weitere Linien, wie die Gattung Hesperocyon (Abb. 6), dem frühesten Vertreter der Hundeartigen, dessen Merkmale aber noch so ursprünglich waren, dass er nur wenig von den Vertretern der frühesten Katzenartigen wie Proailurus und Pseudaelurus (Abb. 7) nur wenig zu unterscheiden ist. Gattungen wie Hemicyon (Abb. 8), bei oberflächlicher Betrachtung einem Hund ähnlich, führte zu den heutigen Bären. Für uns wirken diese Gruppen einander recht ähnlich und der Laie würde sie für sowas ähnliches wie eine Wieselkatze halten, doch die genauere Kenntnis dieser Fossilien belegt, dass sie eben nicht dasselbe waren, sondern evolutionäre Linien, deren Vertreter über viele weitere Abspaltungen zu den heutigen Hunden und Katzen führten (Abb. 9).

Abb. 6: Hesperocyon
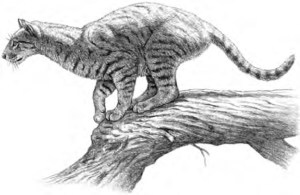

Abb. 7: Proailurus (links) und Pseudaelurus (rechts)

Abb. 8: Hemicyon
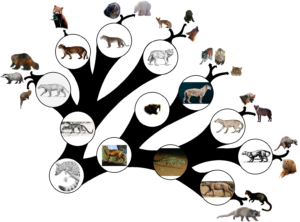
Abb. 9: Evolution der Carnivora
Kreationisten fordern aber, dass sich die neuen Arten so stark von ihren Eltern unterscheiden, dass man nicht einmal sagen kann, dass sie verwandt sind. Die Ironie hierbei ist, dass die Evolutionstheorie niemals besagt, dass eine „Art“ von Dingen jemals zu einer anderen, grundlegend anderen „Art“ von irgendetwas geworden ist, es sei denn, Sie ignorieren alle Zwischenstufen – was natürlich Kreationisten tun.
Die phylogenetische Systematik rekonstruiert die verwandtschaftlichen Beziehungen zwischen den Organismen. Der Evolutionsprozess ist allerdings kleinschrittig. Die einzige Möglichkeit, alle Arten von Leben objektiv zu kategorisieren, besteht in ihren gemeinsamen Merkmalen, die jedes Mitglied dieses Kollektivs und zwar nur dieses Kollektiv hat. Auf diese Weise werden ihre Merkmale diagnostisch und weisen direkt auf eindeutige Gruppen hin.
Um die Evolutionstheorie zu verstehen, muss man nachvollziehen, dass es immer nur darum geht, vorhandene Merkmale und Funktionen zu ändern oder zu verbessern, um auf dem aufzubauen, was bereits vorhanden ist. Entwicklungsbiologie, Genetik und vergleichende Morphologie bestätigen zusammen viele dieser taxonomischen Stadien, so dass Organe und anderen Merkmale nicht plötzlich oder vollständig wie aus dem Nichts erscheinen, da in jedem Fall ein evolutionärer Ursprung erkennbar ist, auf dem sich diese Veränderungen aufgebaut haben.
Sogar der Übergang von Fischen zu Landwirbeltieren, Dinosauriern zu Vögeln oder Affen zu Menschen ist nur eine Frage zunehmender, oberflächlicher Veränderungen, die langsam auf aufeinanderfolgenden Ebenen grundlegender Ähnlichkeiten zusammengestellt werden. Diese stellen monophyletische Gruppen dar, die für immer alle Nachkommen dieser Klade umfassen werden.
Um dies zu verdeutlichen: Wir Menschen sind immer noch Eukaryonten (also Lebewesen, deren Zellen ein Zellkern haben) Wirbeltiere, Säugetiere, Primaten, sind Menschenaffen und eben Menschen.
Mono-Para- und Polyphylie
Wenn wir uns ein Kladogramm weiter anschauen fallen weitere Dinge auf.
Die Verzweigungen werden nicht gewichtet, man hat also kein Maß für die Änderung, um es in einem Kladogramm darzustellen. Alle untersuchten Gruppen (allgemein spricht man von Taxa) stehen am Ende der Verzweigungen. Es wird Auskunft darüber gegeben wie nah die verschiedenen Taxa miteinander verwandt sind. Bei unserem Kladogramm der Säugetiere gehen Beuteltiere und Plazentatiere auf einen gemeinsamen Vorfahren zurück. Sie geben keine Auskunft darüber, welche Gruppe vorher entstanden ist und sie sagen auch nicht aus, dass Beuteltiere „primitiver“ sind als die Plazentatiere. In unserem Kladogramm stehen die Beuteltiere links, die Plazentatiere rechts. Doch genauso gut könnten sie vertauscht stehen, das würde die Aussage des Kladogramms nicht beeinflussen. Entscheidender ist der Verzweigungspunkt. Weil Beuteltiere und Plazentatiere auf einen gemeinsamen Vorfahren zurückgehen, bilden sie, wie oben erwähnt, eine sogenannte monophyletische Gruppe (Abb. 10). Beide Gruppen – Beutel- und Plazentatiere – sind sogenannte Schwestergruppen.
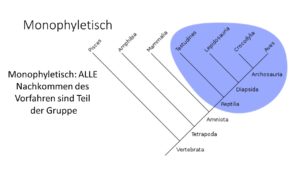
Abb. 10: Monophylie
Monophyletisch wird eine Gruppe im Kladogramm genannt, bei denen alle Vertreter auf einen gemeinsamen Vorfahren zurückgehen. So gehen Beutel- und Plazentatiere alle auf einen gemeinsamen Vorfahren zurück und wie wir in unserem Stammbaum sehen können gehen alle Säugetiere auf einen gemeinsamen Vorfahren zurück. Eine monophyletische Gruppe, auch kurz Monophylum genannt, umfasst alle Untergruppen, die sich aus dem gemeinsamen Vorfahren entwickelt haben, jedoch keine anderen Gruppen.
Untersuchte Gruppen können aber auch paraphyletisch sein (Abb. 11). Das Taxon hat zwar eine jüngste gemeinsame Stammform, enthält aber nicht alle Untergruppen, die auf diese Stammform zurückgehen, wie es beim Monophylum der Fall ist. Sie werden meist durch Plesiomorphien definiert. Ein gutes Beispiel stellt die Klasse der Reptilien dar. Traditionell werden alle kaltblütigen Wirbeltiere mit schuppiger Haut als Reptilien zusammengefasst. Hierzu zählen traditionell die Schildkröten, Echsen, Schlangen, Krokodile, sowie verschiedene ausgestorbene Gruppen wie Dinosaurier. Die Einordnung dieser Gruppen als Reptilien spiegelt jedoch keine evolutionären Verwandtschaftsbeziehungen wider. So sind z. B. die Krokodile mit der Klasse der Vögel (und der ausgestorbenen Dinosaurier) enger verwandt (und bilden das Monophylum der Archosaurier) als mit allen anderen Reptilien. Wenn man also das Taxon Reptilien, also als monophyletische Gruppe, behalten will, muss man die Vögel ebenso zu den Reptilien zählen. Da man dies aber nicht tut, sind Reptilien keine natürliche Evolutionäre Gruppe, sondern ein sogenanntes „wasteback-Gruppen, also paraphyletische Gruppen, die keine evolutionäre Verwandtschaft zeigen. Tatsächlich werden Vögel und die verschiedenen Reptiliengruppen in der Kladistik als Sauropsiden als eine monophyletische Gruppe dargestellt.
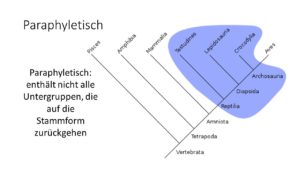
Abb. 11: Paraphylie
Polyphyletisch (Abb. 12) sind Gruppen, die keine gemeinsame Stammform haben und keine natürlichen Gruppen darstellen. Ein Beispiel ist die Warmblütigkeit. Säugetiere und Vögel sind in der Lage ihre Körpertemperatur unabhängig von der Umgebung konstant zu halten. Dieses Merkmal ist aber das Ergebnis konvergenter Evolution, basiert also nicht auf gemeinsamer Abstammung.
Weitere Beispiele für Polyphylie sind solche Bezeichnungen wie “Würmer”, da “Würmer” phylogenetisch unterschiedliche Stämme darstellen, die nur wenig miteinander gemeinsam haben. Auch der Begriff Echse (bzw. Saurier) ist solch eine polyphyletische Bezeichnung, weil viele Gruppen als Echse bezeichnet werden, die nicht eine monophyletische Gruppe bilden.
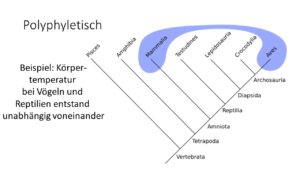
Abb. 12: Polyphylie
Weiterführende Literatur:
BONIK, K. (1981): Evolutionsbiologie und Systematik: Versuch einer Synthese, Verlag Waldemar Kramer, Frankfurt am Main
BURDA, H., HILKEN, G., ZRZAVÝ, J. (2008): Systematische Zoologie. Verlag Eugen Ulmer Stuttgart
FRANZEN, J. L., MOLLENHAUER, D., GUTMANN, W. F. & PETERS, D. S. (1973): Was ist Phylogenetik? Natur und Museum 103 (7): 238 – 242, Frankfurt am Main
Futuyma, D. J. (2007): Evolution. Elsevier, Spektrum, Akademischer Verlag, München
GOULD, S. J. (2002): The structure of evolutionary theory. The Belknap Press of Harvard University Press, Cambridge, MA
Hall, B. (2017): Phylogenetic Trees Made Easy: A How-To Manual. Sinauer; Auflage: 5th ed.
HENNIG, W. (1950): Grundzüge einer Theorie der phylogenetischen Systematik. Deutscher Zentralverlag, Berlin
HENNIG, W. (1966): Phylogenetic Systematics. University Illinois Press, Urbana
Kutschera, U. (2007). Evolutionsbiologie, 2. Auflage. Stuttgart: UTB-Verlag
Neukamm, M. (2001): Die Evolutionstheorie und der moderne Antievolutionismus http://www.martin-neukamm.de/junker.html
Prothero, D. (2017): Evolution – What the Fossils say and why it matters. Second edition. New York: Columbia University Press
Wägele, J. W. (2001): Grundlagen der Phylogenetischen Systematik. Pfeil, F; Auflage: 2
Westheide, W. & Rieger, R. (2007): Spezielle Zoologie (2 Bände). Gustav Fischer Verlag
WIESER, W. (Hrsg.): Die Evolution der Evolutionstheorie Von Darwin zur DNA, Spektrum Akademischer Verlag, Heidelberg, Berlin, Oxford
Williams, D. & Ebach, M. (2020): Cladistics: A Guide to Biological Classification (Systematics Association Special Volume Series, Band 88)Cambridge University Press; Auflage: 3
Wiley, E. O. & Lieberman, B. S. (2011): Phylogenetics: Theory and Practice of Phylogenetic Systematics. Wiley-Blackwell; Auflage: 2
ZRZAVY, STORCH & MIHULKA (2004): Evolution – Ein Lese-Lehrbuch, Spektrum, Akademischer Verlag
