
Die einzige Art alle Arten von Leben objektiv zu kategorisieren ist ihre gemeinsamen Eigenschaften zu erfassen, also diejenigen Merkmale, die jedes Mitglied dieses Kollektivs hat und nur dieses Kollektivs. So werden ihre Merkmale diagnostisch und für diese Gruppe bestimmend. Wir sollten hierbei nicht vergessen, dass der erste Mensch, der versuchte alle Lebewesen zu klassifizieren, ein überzeugter Christ und Kreationist war, der nicht an evolutionäre Prozesse oder eine gemeinsame Abstammung dachte, bzw. davon wusste.
Doch das ursprünglich von ihm entwickelte System, das auch heute noch verwendet wird, bestimmt, dass alles was lebendig ist, in drei Hauptgruppen geteilt werden kann, die sich dann weiter verzweigen in einer fortlaufenden Serie von Unterteilungen. Diese drei Hauptgruppen bezeichnet man als Domänen. Zu diesen gehören die Bacteria, Archaea und Eukarya (Abb. 1).
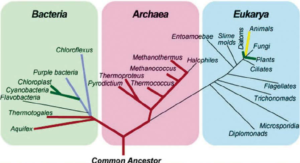
Abb. 1: Die drei Domänen des Lebens
Eukarya oder auch Eukaryoten genannt haben Zellen mit einem Zellkern. Wir gehören zu den Eukarya, wie auch Pflanzen, Tiere, Pilze, Algen und Protista.
Die Genauigkeit der Teilung an der Basis der Eukarya wird noch untersucht, da sich diese Gruppen als sehr vielfältig herausstellen. Es sind jedoch keine Spekulationen erforderlich, um festzustellen, dass Menschen definitiv von Eukaryoten abstammen, da nachweislich jede unserer Zellen einen Zellkern hat. Damit unterscheiden sich die Eukaryoten von den Bacteria und Archaea, also jenen Lebewesen die keinen Zellkern haben und allgemein als Bakterien bezeichnet werden. Doch auch diese sind in ihrem Bauplan und Stoffwechsel so divers, dass sie sich nicht als einheitliche Gruppe zusammenfassen lassen.
Eine bemerkenswerte Untergruppe von Eukarya sind die Opisthokonta (Abb. 2), deren Geschlechtszellen ein einzelnes dorsales Flagellum haben.
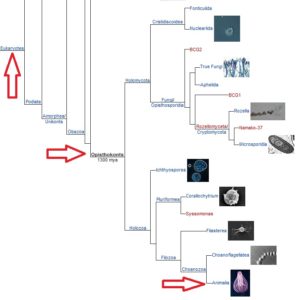
Abb. 2: Stammbaum zu den Opisthokonta
Eine Untergruppe der Opistokonta sind die Metazoa, auch bekannt als das Reich der Tiere (Animalia), mehrzellige Opisthokonten, die andere Lebewesen in einer Art Verdauungstrakt aufnehmen müssen, um zu überleben.
Die biologische Definition und sogar die gängigen Wörterbuchdefinitionen beschreiben den Menschen als zum Tierreich gehörend. Kreationisten heulen bei der Vorstellung, dass sie Tiere sein sollten, aber wenn man weiß, was ein Tier überhaupt ist, dann weiß man, dass wir welche sind! Dies ist auch keine Ansichtssache. Es ist eine Tatsache, und wir können es beweisen! Tiere sind nämlich eukaryotische, mehrzellige Opisthokonta, deren Zellen durch Bindegewebe wie Kollagen zusammengehalten werden und sich von organischem Material ernähren müssen, also heterotroph sind.
Die Taxonomie basiert sowohl auf dem Körperbauplan, der Fortpflanzung und Entwicklung eines Lebewesens als auch auf ihre Körperform selbst. Aus diesem Grund wird das Tierreich dann zwischen den Schwämmen aufgeteilt und alles, was weiter fortgeschritten ist, z. B. die radiärsymmetrischen Nesseltiere und die Bilateria (Abb. 3).
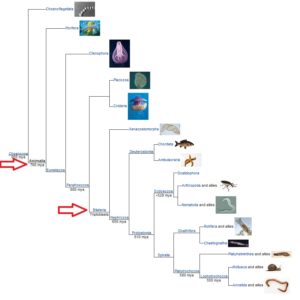
Abb. 3: Bilateria
Bilateria sind Triploblastentiere, das heißt sie entwickeln in einem Stadium ihrer Embryonalentwicklung, der Gastrula, drei sogenannte Keimblätter: Entoderm, Ektoderm und Mesoderm, aus denen sich die verschiedenen Organe eines Lebewesens entwickeln. Ein weiteres wichtiges Merkmal der Bilateria ist ihre Körpersymmetrie: in einem bestimmten Entwicklungsstadium sind diese Tiere bilateral symmetrisch, haben also eine linke und rechte Körperhälfte (Abb. 4). Das trifft übrigens auch auf die radiärsymmetrischen Seesterne zu, da sie von einer Gruppe der Bilateria abstammen und in ihrer Embryonal- und Jugendentwicklung bilateral symmetrisch sind.
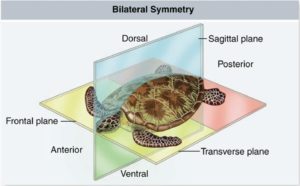
Abb. 4: Bilaterale Symmetrie
Eine der nachfolgenden Unterteilungen sind die Deuterostomia (Abb. 5), Coelomata, bei denen in der Embryonalentwicklung des Darmes der Urmund, also die Mundöffnung des Emybroas, zum After wird und der Mund sekundär aus dem Urdarm durchbricht. Auf gut Deutsch: Deuterostomia beginnen als Arschlöcher…
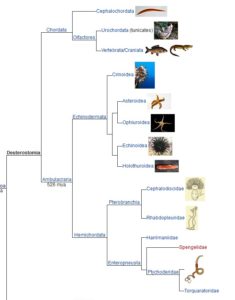
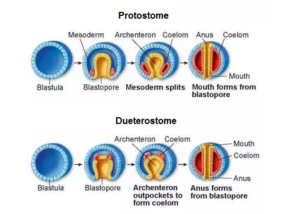
Abb. 5: Deuterostomia
Wir gehören zu den Deuterostomia. Dieses Merkmal teilen wir mit allen Wirbeltieren und weiteren Gruppen wie Seesternen, Seeigeln und Eichelwürmern (Abb. 6). Alles, was wir in der Natur sehen, hält konsequent an allem fest, was wir von einer Kette ererbter Variationen erwarten würden, die durch blühende Abstammungslinien getragen werden, so wie es auch in diesem Fall der Fall ist. Seesterne, Seeigel, Eichelwürmer und alles, was jemals eine Wirbelsäule hatte, entwickeln zuerst die Öffnung für den Anus. Ist das nicht seltsam? Die gemeinsame Abstammung erklärt diese Tatsache offensichtlich, aber bisher konnte kein potenzieller Kritiker der Evolution jemals eine Erklärung dafür oder für einen der anderen Trends liefern, die wir in der Taxonomie sehen.

Abb. 6: Deuterostomia
Die nächste definitiv bestimmbare Unterteilung umfasst die Chordata (Abb. 7), Deuterostomia mit einem Rückgrat. Zu dieser Gruppen gehören die Craniota, Chordaten mit einem Gehirn, welches in einem Schädel eingeschlossen ist. Eine Untergruppe dieser Gruppe umfasst auch Wirbeltiere, von deren Schädel eine Wirbelsäule ausgeht.
Und die nächste Untergruppe sind die Gnathostomata, Wirbeltiere, die all das und einen Kieferknochen haben.
Die nächste Weggabelung liegt zwischen Gnathostomen, deren Skelette entweder knorpelig oder verkalkt sind. Unsere Gabelung führt uns zu Sarcopterygii, knöchernen Wirbeltiere, die sowohl Lungen als auch Beine haben.

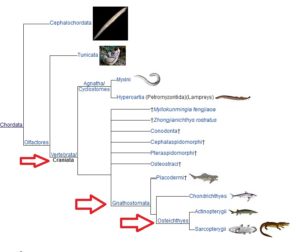
Abb. 6: Chordata, Wirbeltiere, Gnathostomata, Sarcopterygii
Eine Untergruppe davon sind die Stegocephalia (Abb. 7); Wirbeltiere mit Gliedmaßen, an deren Ende sich Finger befinden. Diese Gruppe enthält eine Untergruppe namens Tetrapoda, die jetzt kiemenlose Stegocephalia sind, die vier Gliedmaßen haben. Darin enthalten sind die Reptiliomorpha (Abb. 8), fünffingrige-post-aquatische „terrestrische“ Tetrapoden. Wir sehen jetzt auch eine ausgeprägtere Entwicklung des Gehirns.
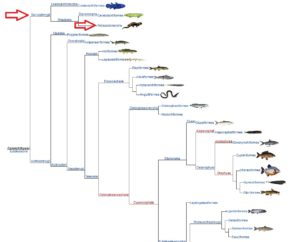
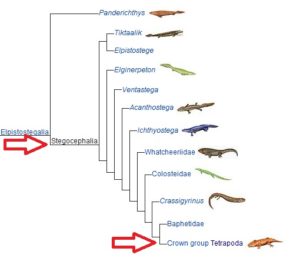
Abb. 7: Stegocephalia
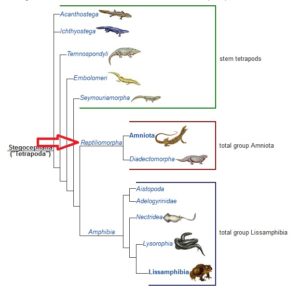
Abb. 8: Reptiliomorpha
Eine der Reptiliomorpha-Untergruppen zeigt eine scheinbar kleine Änderung, die jedoch zu den tiefgreifendsten der Wirbeltierevolution gehört, da der Unterschied entwicklungsbedingt ist. Die Rede ist von der Entwicklung des Amnions. Das Amnion ist eine dünne, durchsichtige, gefäßlose innere Eihaut, die das Embyro umhüllt und so dessen Entwicklung auch außerhalb vom Wasser ermöglicht. Die Entwicklung des Amnions ermöglichte den vollständigen Übergang vom Meer ans Land und wurde offensichtlich von allen Säugetieren, Reptilien und Vögeln geerbt, die seitdem kommen werden (Abb. 9).
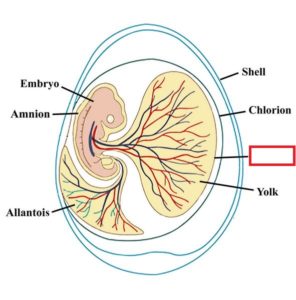
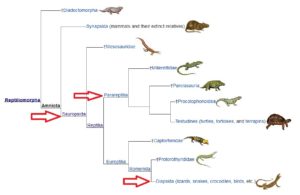
Abb. 9: Amniota
Innerhalb der Amniota vollziehen sich weitere Verzweigungen, die sich u. a. darin unterscheiden wie viele Löcher, sogenannte Schläfenfenster, im Schädel vorkommen. Auf der einen Seite haben wir die Saurposiden, die sich wiederrum in die „Parareptilia“, Amnioten ohne Schläfenfenster und Diapsida, Amnioten mit zwei Schläfenfenstern, einteilen. Die Diapsiden teilen sich wieder ein in die Lepidosauromorpha (Abb. 10), zu denen u. a. die Plesiosaurier und die Schuppenkriechtiere (also Echsen und Schlangen) gehören. Der zweite Ast der Diapsiden sind die Archosaurier, wie sich wiederrum u. a. in Krokodile und die Ornithodira teilen. Zu letzteren gehören die Flugsaurier und die Dinosaurier. Vögel sind dabei eine Untergruppe der Dinosaurier (Abb. 11).
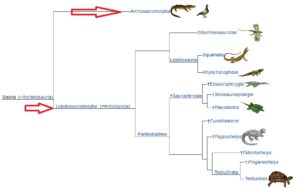
Abb. 10: Sauropsia, Lepidosauromorpha
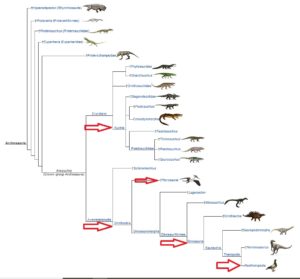
Abb. 11: Sauropsida, Archosauria
Unsere Stammline der Amniota führt zu den Synapsida (Abb. 12), Amnioten mit nur einem Schläfenfenster. Früher wurden sie irrtümlicherweise als säugetierähnliche Reptilien zusammengefasst, eine Bezeichnung, die keiner systematischen Kategorie entspricht. Eine Untergruppe der Synapsida sind die Therapsiden mit zunehmend säugetierähnlichen Skelettformationen sowie rudimentären Stadien, die sich in der kontinuierlich fortschreitenden Entwicklung des Gehirns bemerkbar machen.
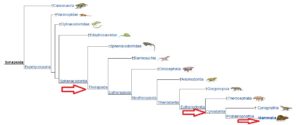
Abb. 12: Synapsida
Innerhalb dieser Gruppe befinden sich die Cynodonten, Therapsiden mit Eckzähnen. Sie sind tatsächlich eine Elterngruppe der Säugetiere, die selbst als endotherme warmblütige Therapsiden mit Milchdrüsen identifiziert wurden. Sogar diejenigen, die schließlich ihre Eckzähne verloren haben, gehören immer noch zu dieser Gruppe, da einige Mitglieder von normalerweise eckzahnlosen Säugetieren immer noch diese Zähne entwicklungsbiologisch, rudimentär oder atavistisch haben. Denn selbst wenn etwas ohne eine oder mehrere diagnostische Merkmale seiner Eltern geboren wird, muss es dennoch als Teil dieser Familie anerkannt werden. Menschen haben bekanntermaßen zwei Arme und zwei Beine. Doch sind jene Kinder ohne Gliedmaßen, die von Eltern mit vier Gliedmaßen geboren wurden, immer noch die Kinder dieser Eltern.
Alle heute lebenden Säugetiere gehören zu einer von drei Hauptabteilungen, die nur einen Bruchteil der wichtigsten Säugetierformen ausmachen, die früher existierten. In mancher Hinsicht ist das Schnabeltier der einzige überlebende Karyotyp, der die Norm der Säugetiervielfalt veranschaulicht, die heute nur noch im Fossilienbestand zu finden ist.
Die bekannteste Gruppe moderner Säugetiere sind die Eutheria, die Brustwarzen wie Beuteltiere haben, aber in einer Plazenta geboren werden, geläufig werden sie auch Placentalia genannt. Diese Gruppe enthält viele Untergruppen auf verschiedenen Ebenen, von denen eine die Gruppen für Primaten enthält. In dieser Gruppe befinden auch uns wir Menschen (Abb. 13).
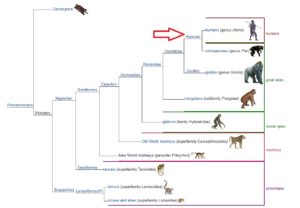
Ich möchte hier in Erinnerung rufen, dass ich entlang des Stammbaums des Menschen nur jener Linie gefolgt bin, die zu uns Menschen führt und jegliche anderen Abzweigungen, die sich teilweise durch eine noch größere Vielfalt auszeichnen, ausgelassen habe, so wie auch nicht jede Zwischenkategorie erwähnt habe. Lediglich hier und da habe ich die ein oder andere Nebenlinie, wie die Diapsiden, angerissen. Hier wird aber schon deutlich, dass im Sinne der phylogentischen Systematik, will man die evolutionären Abstammungslinien nachvollziehen, Linnes 7 Kategorien bei weitem nicht ausreichend sind, Lebewesen taxonomisch einzuordnen. Tatsächlich, würde man die evolutionär-taxonomische Hierarchie vom Reich der Eukarya bis zur Art Homo sapiens durchgehen bräuchte man über 40 taxonomische Kategorien, nach dem heutigen Stand der Taxonomie.
Man sollte immer daran denken, dass wir objektiv beweisen können, dass jedes Mitglied all dieser Gruppen immer noch zu jeder übergeordneten Kategorie gehört, die schon erwähnt wurde.
Z. B. werden Primaten kollektiv definiert als kiemenlose, RNA/DNA-Proteinbasierte, metabolische, metazoische, zellkernenthaltende, diploide, bilateralsymmetrische, endotherme, verdauende, triploblastische, opisthochonte, cölomate Deuterostomia, mit einem Rückgrat, 12 Hirnnerven, die mit einem limbischen System verbunden sind an einer vergrößerten Großhirnrinde mit einem reduzierten Riechfeld in einem Schädel mit Kiefern und spezialisierten Zähnen, inklusive Eck- und Backenzähnen, Vorwärts gerichtetem, voll umschlossenen Augenhöhlen, einem einzelnen Schläfenfenster, befestigt an einem Skelett mit Wirbelsäule, welches vierbeinig und Hinterbein-dominant ist mit einem Becken mit Kreuzbein, Schlüsselbeinen, Handgelenk- und Fußgelenkknochen, die Lungen, Tränendrüsen, Haarfollikel am ganzen Körper, Milchdrüsen, einen opponierbaren Daumen, Nägel und eine verhornte Haut an allen Fingern und Zehen haben, deren Embryonen von einer Amnionhaut umgeben sind, von einer Placenta versorgt werden, lebendgebärend sind und ein komplexes Sozialverhalten haben.
Alle diese Merkmale definieren auch uns als Art Mensch und es ist daher nicht zu leugnen, dass wir aufgrund dieser Merkmale Primaten sind. Wir stammen also nicht vom Affen ab, wir sind Affen. Und wir glauben es nicht, weil wir es glauben wollen, sondern weil wir es beweisen können, es also wahr ist. Menschen wurden seit dem 18. Jahrhundert als Primaten eingestuft, als ein christlicher kreationistischer Wissenschaftler herausfand, was ein Primat war – und andere Wissenschaftler dazu veranlasste, herauszufinden, warum dies auf uns zutraf.
Dem wäre nicht so, wenn verschiedene „Grundtypen“ (von englischsprachigen Kreationisten gerne als „kind“ bezeichnet) des Lebens auf magische Weise erschaffen worden wären, die nicht miteinander oder mit irgendetwas verwandt wären. Es sei denn Gott wollte uns täuschen und dazu bringen zu glauben, dass sich alles entwickelt hat.
Denn der phylogenetische Baum des Lebens ist für jeden objektiven Beobachter, der es wagt, die Anatomie verschiedener Gruppen kollektiver Lebensformen zu vergleichen, von Grund auf klar erkennbar. Aber es kann genauso objektiv von oben nach unten bestätigt werden, wenn es genetisch überprüft wird.
Aus diesem Grund wird es als „doppelt verschachtelte Hierarchie“ bezeichnet. Aber es gibt noch mehr als das, denn die offensichtliche Entwicklung der Physiologie und Morphologie kann sowohl biochemisch als auch chronologisch in der Geologie und entwicklungsbedingt in der Embryologie bestätigt werden. Warum sollte das so sein? Und wie erklären Kreationisten, warum jedes Lebewesen in all diese Tochtergruppen innerhalb der Elterngruppen passt, wobei jede nach scheinbar vererbten Merkmalen abgeleitet wird? Sie versuchen nicht einmal, irgendetwas davon oder irgendetwas anderes zu erklären. Sie werden es nicht tun, weil sie es nicht können, weil die Evolution die einzige Erklärung ist, die all das erklärt.
Weiterführende Literatur:
Phylogeny Explorer https://phylogenyexplorerproject.com/
Phylogeny Research Project http://phylogeny.life/
Tree of Life Web Project http://tolweb.org/tree/
Aron Ra: Foundational Falsehoods of Creationism
BONIK, K. (1981): Evolutionsbiologie und Systematik: Versuch einer Synthese, Verlag Waldemar Kramer, Frankfurt am Main
BURDA, H., HILKEN, G., ZRZAVÝ, J. (2008): Systematische Zoologie. Verlag Eugen Ulmer Stuttgart
FRANZEN, J. L., MOLLENHAUER, D., GUTMANN, W. F. & PETERS, D. S. (1973): Was ist Phylogenetik? Natur und Museum 103 (7): 238 – 242, Frankfurt am Main
Futuyma, D. J. (2007): Evolution. Elsevier, Spektrum, Akademischer Verlag, München
GOULD, S. J. (2002): The structure of evolutionary theory. The Belknap Press of Harvard University Press, Cambridge, MA
Hall, B. (2017): Phylogenetic Trees Made Easy: A How-To Manual. Sinauer; Auflage: 5th ed.
HENNIG, W. (1950): Grundzüge einer Theorie der phylogenetischen Systematik. Deutscher Zentralverlag, Berlin
HENNIG, W. (1966): Phylogenetic Systematics. University Illinois Press, Urbana
Kutschera, U. (2007). Evolutionsbiologie, 2. Auflage. Stuttgart: UTB-Verlag
Prothero, D. (2017): Evolution – What the Fossils say and why it matters. Second edition. New York: Columbia University Press
Wägele, J. W. (2001): Grundlagen der Phylogenetischen Systematik. Pfeil, F; Auflage: 2
Westheide, W. & Rieger, R. (2007): Spezielle Zoologie (2 Bände). Gustav Fischer Verlag
WIESER, W. (Hrsg.): Die Evolution der Evolutionstheorie Von Darwin zur DNA, Spektrum Akademischer Verlag, Heidelberg, Berlin, Oxford
Williams, D. & Ebach, M. (2020): Cladistics: A Guide to Biological Classification (Systematics Association Special Volume Series, Band 88)Cambridge University Press; Auflage: 3
Wiley, E. O. & Lieberman, B. S. (2011): Phylogenetics: Theory and Practice of Phylogenetic Systematics. Wiley-Blackwell; Auflage: 2
ZRZAVY, STORCH & MIHULKA (2004): Evolution – Ein Lese-Lehrbuch, Spektrum, Akademischer Verlag
