
Lesetext: Homologie
In den vorherigen Kapiteln haben wir uns intensiv mit der phylogenetischen Systematik und Kladistik befasst.
Wir halten fest: Lebewesen sind ein Mosaik aus apomorphen und plesiomorphen Merkmalen. Doch wie bestimme ich, welche Merkmale apomorph und welche plesiomorph sind?
WAS IST HOMOLOGIE?
Hierbei sind u. a. Homologie-Kriterien hilfreich. Gehen die Merkmale auf einen gemeinsamen evolutionären Ursprung zurück, spricht man von Homologien.
Im Wesentlichen sind drei Homologie-Kriterien bekannt (REMANE 1952, vgl. auch FUTUYMA 2007, WIESER 1994, PROTHERO 2017)
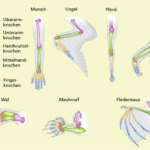
1. das Kriterium der Lage, Dieses sagt aus, dass Strukturen zueinander homolog sind, wenn sie trotz unterschiedlicher Ausprägung in Gestalt und Anzahl in einem vergleichbaren Gefügesystem stets die gleiche Lagebeziehung aufweisen. Ein bekanntes Beispiel hierfür sind die Vordergliedmaßen der Wirbeltiere (Abb. 1). Egal ob wir einen Menschen, ein Pferd, einen Vogel oder eine Echse haben, so ist der Grundaufbau der Vordergliedmaßen vom Prinzip her gleich: ein Oberarmknochen, zwei Unterarmknochen, mehrere Handwurzelknochen, bis zu fünf Mittelhandknochen und bis zu fünf Finger. Trotz des gleichen Grundaufbaus kam es im Verlauf der Erdgeschichte zu verschiedenen Umwandlungen, da die Gliedmaßen unterschiedliche Zwecke erfüllen: laufen, graben, schwimmen, fliegen etc. Einzelne Knochen verschmolzen oder wurden reduziert, die Proportionen änderten sich, doch der Grundaufbau bleibt der gleiche und dieser weist auf gemeinsame Vorfahren hin, von denen aus sich die Entwicklung zu den heutigen Formen vollzogen hat.
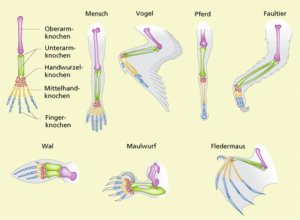
Abb. 1: Kriterium der Lage
2. das Kriterium der spezifischen Qualität und Struktur, welches besagt, dass Strukturen dann zueinander homolog sind, wenn sie einen übereinstimmenden Feinbau besitzen. Ein bekanntes Beispiel hierfür ist der Vergleich unserer Zähne mit den Haifisch-Schuppen (sog. Placoidschuppen, Abb. 2). Beide haben denselben Feinbau: Die oberste Schicht besteht aus Zahnschmelz und darunter befindet sich das Dentin. Die Gemeinsamkeit des Aufbaus lässt sich damit begründen, dass evolutionsgeschichtlich die urtümlichen kieferlosen Fische über ein Außenskelett verfügten, welches aus Placoidschuppen bestand. Einige dieser Schuppen im Bereich der Mundöffnung hatten sich für die Nahrungsaufnahme spezialisiert und wurden zu Zähnen. Mit der Ausbildung einer knöchernen Wirbelsäule hatte ein stabiles Außenskelett keine Notwendigkeit mehr und wurde reduziert. Übrig geblieben sind lediglich die Zähne und bei den Haien zusätzlich die Placoidschuppen (die eine wichtige Funktion einer effizienten Schwimmbewegung übernehmen).
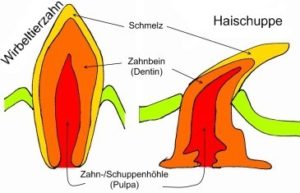
Abb. 2: Kriterium der spezifischen Qualität und Struktur
3. Das dritte Homologiekriterium ist das Kriterium der Kontinuität. Strukturen sind dann zueinander homolog, wenn sie durch eine Reihe gleichartiger Übergangsformen miteinander verbunden sind. Beispiele für das Kriterium der Kontinuität sind z. B. die Kreislaufsysteme der Wirbeltiere oder die Entwicklung der Gehörknöchelchen der Säugetiere, die ursprünglich Teile des Kieferapparates waren (Abb. 3).
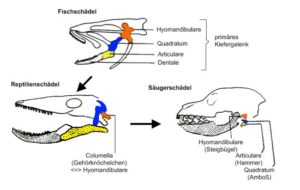
Abb. 3: Kriterium der Kontinuität
Neben morphologischen Daten werden zunehmend molekulare Daten genommen. Molekulare Methoden haben längst einen wichtigen Status in allen biologischen Bereichen erhalten, so auch in der Ermittlung verwandtschaftlicher Verhältnisse und evolutiver Zusammenhänge. Sie haben im Vergleich zu morphologischen Daten einige Vorteile:
Zum einen sind sie leichter zu gewinnen. Zum anderen werden DNA-Moleküle direkt vererbt und unterliegen einer geringeren Beeinflussung durch die Umwelt, während morphologische Merkmale wesentlich variabler und damit plastischer sind. Sie sind in ihrer Ausprägung daher einfacher und evolvieren insgesamt gleichförmiger als morphologische oder physiologische Merkmale. Sie zeichnen sich auch durch gemeinsame Homologien aus: je mehr Gen-Abschnitte sich ähneln, desto höher ist die Verwandtschaft. Man denke nur an den berühmten Vaterschaftstest, der im Prinzip nach demselben Muster funktioniert (und von Kreationisten nicht angezweifelt wird, außer vielleicht bei der Jungfrauengeburt Marias).
Es ist nicht nur so, dass man diese Homologien auf gewisse Ähnlichkeiten zwischen den Lebewesen hindeuten, sondern, dass man anhand dieser Homologien die Lebewesen in ein hierarchisch geschachteltes System gemeinsamer Abstammung zuordnen kann (REMANE et al. 1973, AX 1984, WÄGELE 2000, MAHNER & BUNGE 2000,OSCHE, 1979) . Hierzu gehören auch Kenntnisse des Feinbaus, der Genetik, der Embryonalentwicklung usw. usf. D. h. man untersucht die Merkmale und Homologien der Lebewesen bis ins möglichst letzte Detail.
WAS IST ANALOGIE?
Von Homologien sind die Analogien zu unterscheiden (REMANE 1952, REMANE et al. 1973, SHUBIN 1994, SUDHAUS & REHFELD 1992). Analoge Organe haben dieselbe Funktion, aber keine gemeinsame Abstammung. Wenn wir die Flügel von Fledermaus und Vogel vergleichen, so erkennen wir, dass zwar die Anordnung der Armknochen zueinander homolog ist, die Flügel als Gesamtkonstruktion sind jedoch völlig anders gebaut. Fledermäuse haben besonders lange Finger, um die sich eine Flughaut spannt, während bei Vögeln die Fingerknochen verschmolzen bzw. reduziert sind und sogenannte Armschwingen haben. Zusätzlich haben Vögel Federn (Abb. 4).
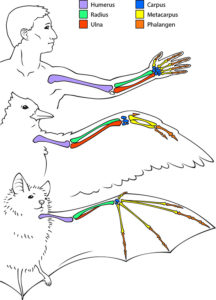
Abb. 4: Vergleich Flügel Vogel – Fledermaus
Aus diesem Vergleich ist zu schlussfolgern, dass Vögel und Fledermäuse aufgrund ihrer Homologie der Anordnung der Armknochen auf einen gemeinsamen Vorfahren zurückzuführen sind, ihre Flugfähigkeit sich aber unabhängig voneinander entwickelt hat. Selbe Knochenstruktur, aber andere – unabhängige Lösungswege den Luftraum zu erobern. Man bezeichnet sowas auch als konvergente Evolution. Konvergente Evolution kommt vor, wenn auf die Lebewesen ein ähnlicher Selektionsdruck wirkt, diese aber unabhängig voneinander Lösungen für diesen Selektionsdruck entwickeln.
VORWÜRFE DER KREATIONISTEN
Kreationisten unterstellen gerne, dass Evolution mit Ähnlichkeit (Homologie) begründet und umgekehrt Homologie durch evolutionstheoretische Annahmen bestimmt werde (LÖNNIG, 1998, JUNKER & SCHERER 2013, WELLS 2000). Damit entstünde, so die Kritiker, ein Zirkel gegenseitiger Selbstbestätigung, wodurch das evolutionstheoretische Homologieargument an Wert verlöre. Dabei können diese Ähnlichkeiten aber auch von einem gemeinsamen intelligenten Designer, also Gott, geschaffen worden sein.
Kreationisten übersehen jedoch, wie perfekt diese Homologien in ein hierarchisch ineinander geschachteltes System gemeinsamer Abstammung passen. Wir schauen uns nicht einfach bloß die Armknochen von Wirbeltieren an, sehen, dass sie dieselbe Knochenstruktur haben und dann einfach sagen, sie haben einen gemeinsamen Vorfahren. Dazu gehört mehr.
Tatsächlich ist es so, dass bevor eine Struktur sicher als Homologie definiert werden kann, muss vorher die gemeinsame Abstammung dieser Strukturen geklärt und begründet werden. Homologien werden also erst definiert, nachdem diese als gesichert für eine gemeinsame Abstammung gelten.
Die Seite talkorigins gibt folgende Erklärung hierzu:
Homologie wird nicht als Ähnlichkeit aufgrund gemeinsamer Abstammung definiert und dann als Beweis für gemeinsame Abstammung verwendet. Der Beweis für eine gemeinsame Abstammung stammt vielmehr aus den Ähnlichkeitsmustern vieler Merkmale. Diese Ähnlichkeiten zeigen, dass sich Organismen auf natürliche Weise in ein hierarchisch verschachteltes System zusammenschließen. Zum Beispiel, dass Marienkäfer und Mistkäfer beides Käferarten sind, basiert auf verschiedenen gemeinsamen Merkmalen wie z. B. gehärteten Vorderflügeln; Käfer, Fliegen und Heuschrecken sind Typen von Insekten; Insekten, Skorpione und Tausendfüßler sind Typen von Arthropoden. Eine solche Gruppierung hängt nicht von Annahmen über die Herkunft ab und wurde tatsächlich zuerst von Linne, einem Kreationisten, kodifiziert. Eine Gruppierung, die von vielen gemeinsamen Merkmalen vorgeschlagen wird, ist ein Beweis für eine gemeinsame Abstammung. Dies gilt unabhängig davon, wie die Merkmale genannt werden. Das Homologie-Label wird hinzugefügt, nachdem die Beweise für gemeinsame Vorfahren bereits vorliegen. Quelle
Das heißt homologe Strukturen werden erst als solche definiert, nachdem wir genügend Beweise gesammelt haben, dass diese von einem gemeinsamen Vorfahren abstammen (vgl. REMANE et al. 1973, AX 1984, WÄGELE 2000, MAHNER & BUNGE 2000, OSCHE 1979 und MINDEL & MEYER 2001, LAUDER 1994) .
HOMOLOGIE ODER DESIGN?
Treffen aber solche Beobachtungen und Homologietheorien auch für einen intelligenten Designer zu? Könnte man nicht einwenden, dass diese Ähnlichkeiten der Lebewesen auf einen gemeinsamen Schöpfer zurückgehen?
Wenn es irgendeinen intelligenten Designer gäbe, der alles auf einmal erschaffen hätte, gäbe es keinen Grund, dass er sich an diese Homologiekriterien halten müsste.
So könnte er Fische mit Haaren produzieren, er könnte ein Säugetier erschaffen, welches vier Beine und zwei Paar Flügel hätte und kein Wirbeltier ist oder einen Käfer, der nicht gleichzeitig ein Insekt ist. Natürlich könnte man einwenden, dass ein intelligenter Designer, der dieselben Materialien nutzt, homologe Ähnlichkeiten erschaffen haben könnte. Aber bei der enormen Vielfalt an Lebewesen und der unterschiedlichen Anpassungsstrategien ist es unwahrscheinlich, dass ein intelligenter Designer dahintersteckt. Denn bei jedem erdenklichen Beispiel lässt sich die Homologie, also die gemeinsame evolutionäre Abstammung, beweisen.
Man vergleiche z. B. Fische und Delfine miteinander: beide Bautypen haben eine ähnliche Anpassung: einen stromlinienförmigen Körper, um im Wasser zu schwimmen. Aber dennoch ist diese Körperform keine Homologie, sie entstand unabhängig voneinander (Vgl. Abb. 5). Denn wenn wir die Anatomie dieser Tiere genauer untersuchen, merken wir z. B., dass Delfine den Körperbauplan eines Säugetieres haben, ihr Körperbau also homolog zu denen der anderen Säugetiere ist. Delfine haben z. B. Lungen, die dem Bau der Lungen von Säugetieren homolog sind. Warum stattete der intelligente Designer aber Delfine nicht mit einem zusätzlichen Paar Kiemen aus, wie Fische sie haben, wenn er Delfinen schon eine ähnliche Fischform gab? Wenn der Designer doch alle Tiere gleichzeitig erschaffen hat, warum verwendete er das Fischmaterial nicht für die Delfine? Oder warum machte er Delfine nicht gleich zu Fischen? Litt der intelligente Designer unter Mangelwirtschaft? Ging das Klopapier zuneige? Warum dachte er sich dann überhaupt sowas wie Delfine aus und wenn ihm Material fehlte, scheint er zumindest nicht allmächtig gewesen zu sein. Außerdem: Lungenfische haben z B. auch beides: Kiemen und Lungen, sind aber trotzdem weder Delfine noch Säugetiere.

Abb. 5: Konvergenz
Delfine passen innerhalb des hierarchischen Systems der Taxonomie und phylogenetischen Systematik zweifelsfrei in die höhere Kategorie der Säugetiere und nicht der Fische.
Und alles was in die Säugetierkategorie passt, passt auch in die Amnioten-, Tetrapoden- und Wirbeltierkategorie. Amnioten definieren sich dadurch, dass sich um das Embryo die Amnionhülle entwickelt, die die Embryonalentwicklung unabhängig vom Wasser macht. Das amniotische Ei – oder als Neuentwicklung bei der Säugetier- Unterkategorie der Plazentatiere das austragen des Embryos im Mutterleib – ist typisch für alle Säugetiere und Sauropsiden (dazu gehören Reptilien und Vögel). Alle Amnioten sind auch Tetrapoden (also alle Landwirbeltiere mit ursprünglich vier Gliedmaßen) zu denen sich neben den Amnioten dann zusätzlich Amphibien dazu gesellen. Und all diese Tetrapoden gesellen sich zusammen mit den verschiedenen Gruppen, die wir als „Fische“ bezeichnen, zu den Wirbeltieren.
Amphibien haben zwar gemeinsamen Merkmale mit allen anderen Tetrapoden und Wirbeltieren (z. B. das Vorhandensein einer Wirbelsäule und vier Gliedmaßen), jedoch nicht mit den Amnioten, weil Amnioten neben den typischen Tetraopdenmerkmalen weitere Merkmale besitzen, die Amphibien nicht haben (Amphibien legen z. B. kein amniotisches Ei) – und Amphibien umgekehrt Merkmale haben, die den Amnioten fehlen. Damit haben Amphibien auch keine Säugetiermerkmale, da sie weder ein Fell haben, noch Milch produzieren und Säugetiere keine Kaulquappen.
Würde ein intelligenter Designer mit denselben Materialien arbeiten, müsse man sich die Frage stellen, warum Amphibien zwar Merkmale haben, die allen Wirbeltieren gemeinsam sind, aber keine Merkmale, die typisch für Säugetiere sind, obwohl Säugetiere ja auch Wirbeltiere sind?
Warum sollte ich Lebewesen mit denselben „Materialien“ erstellen und dann bei ausgewählten Tiergruppen zusätzliche Merkmale entwickeln, bei einigen auslassen etc.?
Und wenn Amphibien laut Aussage der Kreationisten etwas vollkommen anderes sind als Säugetiere, warum haben die dann Merkmale, die Amphibien zusammen mit Säugetieren nebst anderen Klassen als Wirbeltiere zusammenfassen?
Dasselbe gilt im Übrigen auch für uns: Laut kreationistischer Behauptung sind wir ja etwas völlig anderes als Tiere. Gott erschuf ja schließlich den Menschen separat. Aber warum finden wir dann so viele gemeinsame Merkmale mit anderen Affenarten, dass wir quasi gesehen in dieselbe Gruppe eingeordnet werden? Selbst Carl von Linne, den wir in einem anderen Video kennengelernt haben, ordnete uns zu den Primaten, obwohl er von Evolution keinen Schimmer hatte und von der Unveränderlichkeit der Arten ausging. Wären wir Menschen eine völlig andere Schöpfung als die der Tiere, die uns so sonderbar machen, warum haben wir nicht einen Bauplan, der solche Homologien gar nicht erkennen lassen würde: Warum haben wir, wie alle Tiere, Zellen mit einem Zellkern? Warum haben wir eine Wirbelsäule, wie alle Wirbeltiere? Warum haben wir nicht ein extra Paar Beine, um uns von den Tetrapoden zu unterscheiden? Warum ist unsere DNA mit den Schimpansen ähnlicher, als die DNA der Schimpansen mit denen der Gorillas, wenn wir etwas völlig anderes sind, als ein Affe? Wieso akzeptieren Kreationisten einen DNA-Test unter Familienmitgliedern als definitive Verwandtschaft, lehnen aber dieselbe Methode für evolutionäre Verwandtschaften mit anderen Arten ab?
Nicht zu vergessen: homologe Merkmale sind auch Merkmale, die an die nächste Generation vererbt werden. Durch die Vererbung wissen wir, dass Merkmale einen gemeinsamen Ursprung haben.
Und wir wissen, dass diese vererbten Merkmale variieren. So wie Kinder ihren Eltern ähneln und diese ihren anderen Familienmitgliedern und sich so zu größeren Familienstammbäumen gruppieren ist es genauso mit den Arten. Ähnliche Arten haben ihre Merkmale von anderen ähnlichen Vorläufer-Arten geerbt etc., mit denen sie sich zu größeren Einheiten gruppieren und die verschiedenen systematischen Einheiten bilden.
Dass strukturelle „Information“ genetisch an die Nachkommen vererbt wird, ist einleuchtend. Die Vererbung stellt, salopp formuliert, ein „Gedächtnis“ dar, „einen Speicher für alle Erfolge, die das Leben jemals errungen hat.“. Wenn also die Merkmale von Generation zu Generation weitervererbt werden, wenn zugleich aber auch ein allmählicher Wandel der Arten stattfindet, dann ergibt sich die logische Folgerung, dass zwischen den Arten eine (abgestufte) Formenähnlichkeit bestehen muss, die sich vom morphologischen bis hinab zum molekularen Bereich erstreckt. So ist für uns die Abstammung offensichtlich, wenn ein Kind seinen Eltern „wie aus dem Gesicht geschnitten“ oder der Enkel in seinem Verhalten „ganz der Opa“ ist.
Entsprechend kann auch an der Verwandtschaft zwischen den verschiedenen Arten kein rational begründeter Zweifel bestehen, wenn man hier auf tiefgreifende Formenähnlichkeiten stößt – vorausgesetzt, man erkennt die Standards wissenschaftlichen Argumentierens an. Vergleicht man die Organismen verschiedener Arten und Gruppen miteinander, stellt man tatsächlich fest, dass zwischen ihnen mehr oder minder große Ähnlichkeiten existieren. Dies wurde z. B. an den Vordergliedmaßen der Wirbeltiere verdeutlicht, die bei den Arten zum Teil völlig unterschiedliche Funktionen übernehmen und dennoch einen fast identischen Feinbau besitzen (REMANE et al.1973, OSCHE 1979, GOULD 1987, BERCK 2002, NEUKAMM 2002, AX 1984, WÄGELE 2000).
Das weist darauf hin, dass gerade solch komplexe Merkmalsgruppierungen, die bei verschiedenen Arten detailgetreu in Erscheinung treten, aber im Dienst verschiedener Funktionen stehen, auf Abstammung von einem Vorfahren hindeuten. Wie eine solche Datensituation völlig ohne Vererbung und Evolution erklärt werden könnte, ist meines Erachtens bislang noch nicht überzeugend aufgezeigt worden. Die strukturellen Übereinstimmungen lassen sich in neuerer Zeit bis in den molekularen Bereich hinein verfolgen, wobei sich zeigt, dass (fast) alle rezenten Lebewesen nicht nur denselben genetischen Code besitzen, sondern auch in weiten Teilen über ein nahezu identisches Repertoire an Genen, Biomolekülen und Stoffwechselprozessen verfügen (KUTSCHERA 2007). Wenn sich durch Aufspaltung und Umwandlung von Stammarten schrittweise neue Arten gebildet haben und einmal getroffene „Entscheidungen“, die sich als erfolgreich herausgestellt haben, kaum mehr rückgängig zu machen oder bestenfalls nur sehr eingeschränkt veränderbar sind, müssen sich die vererbten Merkmale auch in eine abgestufte Rangfolge bringen lassen. Daraus resultiert dann im Idealfall eine gesetzmäßige Beziehung zwischen den Merkmalen oder anders ausgedrückt: eine abgestufte Formenähnlichkeit, ein hierarchisches System konsequent ineinander geschachtelter Gruppen.
Die hierarchische Struktur der Taxonomie macht also nur Sinn, wenn man von einer gemeinsamen Abstammung ausgeht.
Diese Hierarchie der systematischen Gruppen ist nun ein Beleg für die Evolutionstheorie, weil das Auftreten gesetzmäßig abgestufter Merkmalsgefüge – in Kombination mit dem Wissen um die Vererbung von Merkmalen, Variation und Selektion – ohne DARWINs Abstammungsthese nicht vernünftig zu erklären ist. (REMANE et al., 1973).
Natürlich, Kreationisten könnten immer noch einwenden, dass dies auf einen gemeinsamen Schöpfer zurückzuführen ist. Dennoch lässt sich das gewünschte Ziel, nämlich die Schöpfungsthese damit wissenschaftlich zu rechtfertigen, nicht erreichen, weil der Gedankengang methodisch fragwürdig ist. Eine solche Behauptung, dass dahinter ein gemeinsamer Schöpfer steckt, enthält kein wissenschaftliches Argument gegen die Evolution, weil praktisch alle Fakten auf diese Weise erschlossen werden, es wird also für alles (und damit nichts) ein Schöpfer postuliert. Man kann dann nur noch die Methode der Naturwissenschaft generell infrage stellen und damit den Boden der wissenschaftlichen Argumentation verlassen. Damit ist der intelligente Designer wissenschaftlich nicht ergründbar. Und genau das ist der springende Punkt: Religion oder Glaube ist eben keine Wissenschaft und daher nicht mit wissenschaftlichen Methoden greifbar. Dadurch würde sich jegliche Diskussion um Schöpfung oder Evolution erübrigen, würden Kreationisten nicht versuchen, ihren Schöpfungsmythen ein wissenschaftliches Antlitz zu verleihen.
Die Schöpfungshypothese ist keine wissenschaftliche Alternative zur Evolutionstheorie. Denn was der „Designer“ erschaffen hat, wie und warum er das alles so und nicht anders erschuf, wie er es getan hat, kann man mit den „natürlichen“ Mitteln, die uns zur Verfügung stehen, prinzipiell nie erforschen. Der Schöpfer braucht sich weder zu erkennen geben, noch muss er sich innerweltlichen Gesetzen beugen oder mit Materie wechselwirken. Der „intelligente Designer“ bleibt von vorn herein ein unlösbares Mysterium und erklärt daher nichts. Wenn in der Evolutionsbiologie zahlreiche Fragen unbeantwortet geblieben sind, bedeutet das nicht, dass wir die Antworten prinzipiell nie finden können, sondern eben nur, dass wir vor praktischen Problemen der Erkenntnis stehen.
Ein Biologe kann auf naturalistischer Grundlage weiterforschen, und es besteht die berechtigte Hoffnung, dass wir im Laufe der Zeit immer mehr offene Fragen beantworten können. Dahingegen bringt die Forschung die Schöpfungstheorie nicht weiter, weil wir prinzipiell nicht wissen können, was sich der „Designer“ wohl gedacht haben mag.
PROBLEME DER HOMOLOGIE
Natürlich treten bei der Konstruktion von Kladogrammen aufgrund diverser Homologien einige Schwierigkeiten auf, die von Evolutionsgegnern mit Vorliebe zur Kritik an der Evolutionstheorie genutzt werden. So kann man zunächst oft nicht eindeutig beurteilen, welche abgeleiteten Merkmale abstammungsbedingte Homologien sind und welche Merkmale unabhängig voneinander entstanden sind (SHUBIN 1994, SUDHAUS & REHFELD 1992).
Dieses Problem löst man damit, indem mehrere Fälle durchgespielt werden, d. h. unterschiedliche Kladogramme entwickelt werden, die die möglichen Evolutionsszenarien aufzeigen und man gegeneinander abwägt. In der Kladistik sind nun Kladogramme am plausibelsten, das heißt am „sparsamsten“, wenn die Zahl an Apomorphien maximal und die Zahl der Konvergenzen minimal ist. Man spricht auch von der Maximum Parsimony (RIEPPEL, 1999, S. 38; WÄGELE 2000). Das macht insofern Sinn, als dass Evolution ein langsamer Prozess ist und Veränderungen daher sich langsam ausbreiten. Bei der Beurteilung der Plausibilität eines Kladogramms spielt jedoch auch die Güte der abgeleiteten Merkmale eine Rolle, die je nach Komplexität gewichtet werden müssen.
Es gibt aber eine Reihe von Lebewesen, die nicht außerordentlich gut erforscht sind – was bei über 2 Mio. lebenden Arten nun auch nicht verwundern sollte. Daher sind Homologien nicht bei jedem so einfach festzustellen oder von Konvergenzen abzuleiten.
So ist zwar die Phylogenie der Wirbeltiere recht gut erforscht, doch bei allem, was man gerne als „Gewürm“ und „Krabbelviecher“ abwertend bezeichnet, ist die Sache etwas komplizierter. Entsprechend gibt es bei vielen Gruppen unterschiedliche Hypothesen eines Stammbaums.
Ein gutes Beispiel ist z. B. das der Verwandtschaft innerhalb der Gruppe der Euarthropoda. Zu diesen gehören die Insekten (Hexapoda), Spinnen (Chelicerata), Krebstiere (Crustacea) und Tausendfüßer (Myriapoda). Die Gruppe der Euarthropoda ist die artenreichste Tiergruppe überhaupt, mit etwa 1,5 Mio. beschriebenen Arten. Bei so vielen Arten sind die Merkmale, Homologien und Konvergenzen so komplex, dass unterschiedliche Verwandtschaftshypothesen postuliert werden (Vgl. BURDA et al. 2008).
Keineswegs ist aber diese Stammbaumvielfalt so zu interpretieren, dass man keine Ahnung habe und die Evolutionstheorie zu einer unfruchtbaren und wilden Spekulation verkomme. Nun wird kein Evolutionsbiologe bestreiten, dass Probleme in der Stammbaumkonstruktion bestehen. Es wäre jedoch verkehrt, daraus die Falschheit des Evolutionsgedankens abzuleiten:
Erstens kann man das Problem der „Stammbaumvielfalt“ minimieren, indem man einfach soviele Merkmale wie möglich zur Erstellung von Stammbäumen heranzieht.
Werden Stammbäume nur anhand weniger Merkmale erstellt, sind die Resultate in der Tat oft widersprüchlich – zudem ist bei einer geringen Anzahl der verwendeten Merkmale die Gefahr groß, dass konvergent entstandene Merkmale stärker betont werden (MISHLER et al. 1994)
Zweitens führt keine Analyse zur Feststellung völlig chaotischer Merkmalsverteilungen. Viele Merkmale sind gesetzmäßig miteinander verbunden, eine „freie“ Kombination der Merkmale gibt es nicht. Das heißt man findet im Ganzen eine hierarchische und „von gegenseitiger Abhängigkeit geprägter Ordnung“, die von Konvergenzen zwar gestört aber nicht aufgehoben wird (RIEDL 1990). Stammbäume sind deshalb keine Fiktion, wir sind uns nur oft (noch) nicht im Klaren darüber, welcher Stammbaum „der Richtige“ ist!
Schließlich sollte drittens nicht unerwähnt bleiben, dass beim Vergleich zwischen den Modellstammbäumen der vergleichenden Morphologie und denen der Molekularbiologie oft ein mehr oder minder hoher Grad an Übereinstimmung zutage tritt. Die Daten der Molekularbiologie bestätigen die vergleichende Morphologie in praktisch allen gut abgesicherten Fällen (MAIER 1994). Dort wo Widersprüche zutage treten hat sich entweder die Morphologie getäuscht oder die untersuchten Genabschnitte waren fehlerhaft. Weitere Forschungen haben solche Diskrepanzen zum Teil lösen können.
Es wäre an den intelligenten Designer auch zu fragen, warum er sich die Mühe machen muss, dass genetische Daten mit morphologischen so gut übereinstimmen.
Doch erst im Rahmen des von DARWIN vorgeschlagenen, theoretischen Evolutionsmechanismus (Variation und natürliche Zuchtwahl) ergeben sich eine Reihe logischer Folgerungen aus der Abstammungshypothese, die mit der Beobachtung im Einklang stehen.
Diese These wird auch durch den Fossilienbefund gestützt, der ebenfalls zu den klassischen Belegen für eine gemeinsame Stammesgeschichte der Arten zählt (BENTON 1998, PAGE & COTTON 2002). Weil sich in der Evolution die schrittweise Abänderung von Merkmalen vollzieht, ergibt sich nämlich die Forderung, dass es fossil erhaltene Arten geben muss, die in ihren mosaikartigen Merkmalsgefügen „zwischen“ den heute existierenden Organismengruppen stehen. Betrachten wir die fossil überlieferten Tier- und Pflanzenarten, so stellt man in der geochronologischen Abfolge der Fossilien tatsächlich keinen chaotischen, sondern einen geordneten Formenwandel fest. Die Lebewesen haben sich im Laufe der Zeit in ihrem Bau immer mehr den heutigen Formen angeglichen.
So hat z.B. Michael J. BENTON (1998) in seiner Arbeit „Molecular and Morphological Phylogenies of Mammals: Congruence with Stratigraphic Data“ 206 Kladogramme, die aus morphologischen und molekularen Daten von Säugetieren gewonnen wurden, mit den fossilen Daten verglichen. Im Ergebnis zeigt sich, dass die morphologischen Daten etwas besser mit dem Fossilbericht korrespondieren, als die molekularen Stammbäume. Insgesamt stellt BENTON aber eine gute Übereinstimmung zwischen Systematik und Stratigraphie fest, und zwar sowohl bezüglich der morphologischen als auch hinsichtlich der molekularen Daten.
Die Stammbäume lassen sich also mit der chronologischen Abfolge der „großen Gruppen“ in den geologischen Schichten recht gut in Einklang bringen, das heißt die ranghöheren systematischen Gruppen korrelieren meist mit einem höheren radiometrischen Alter der entsprechenden fossilen Vorfahren. Ja, in einigen Fälle ist es sogar gelungen, den Zeitraum, innerhalb dessen bestimmte Artaufspaltungen stattfanden, durch „molekulare Uhren“ abzuschätzen und das Ergebnis anhand geologischer bzw. biogeographischer Befunde zu untermauern.
Homologien sind nicht einfach nur auf ein oberflächliches Vergleichen von Ähnlichkeiten zurückzuführen, sondern man geht in die tiefere Analyse des Aufbaus der Lebewesen. Dabei ist es bezeichnend, dass jede dieser Analysen, wirklich JEDE, in ein hierarchisch geschachteltes System gemeinsamer Abstammung passt.
Die Beständigkeit der Homologien sind ein Beweis für die gemeinsame Abstammung, weil sie ständig durch neue Daten geprüft wird, die Hypothesen gemeinsamer Abstammung also ständig aktualisiert werden, wobei alte Daten durch neuere ergänzt werden – so wie es in jeder Wissenschaft der Fall ist.
Literatur
AX, P (1984) Das Phylogenetische System. Stuttgart, New York
BENTON, MJ (1998): Molecular & Morphological Phylogenies of Mammals: Congruence with Stratigraphic data. Molecular Phylogenies & Evolution 9 (3), 398-407
BERCK, KH (2002) Evolutionstheorie im Biologieunterricht – ein Vorschlag. Praxis der Naturwissenschaften – Biologie 1/51, S. 35-38
BURDA, H., HILKEN, G., ZRZAVÝ, J. (2008): Systematische Zoologie. Verlag Eugen Ulmer Stuttgart
FUTUYMA, D. J. (2007): Evolution. Elsevier, Spektrum, Akademischer Verlag, München
GOULD, SJ (1987) Der Daumen des Panda. Betrachtungen zur Naturgeschichte. Birkhäuser, Stuttgart
JUNKER, R. & SCHERER, S. (2013, Hrsg.): Evolution: Ein kritisches Lehrbuch. Weyel; Auflage: 7. aktualis. Aufl.
KUTSCHERA, U. (2007). Evolutionsbiologie, 2. Auflage. Stuttgart: UTB-Verlag
LAUDER, GV (1994) Homology, form and funktion. In: HALL BK (Hrsg.) Homology: The hierarchical basis of comparative biology. San Diego, S. 150-196
LÖNNIG, WE (1998) Johann Gregor Mendel – Warum seine Entdeckungen 35 (72) Jahre ignoriert wurden.
MAHNER M & BUNGE M (2000) Philosophische Grundlagen der Biologie, Springer-Verlag, Berlin
MAIER UG (1994) Die molekulare Dimension der biologischen Evolutionstheorie. In: WIESER W (Hrsg.) Die Evolution der Evolutionstheorie. Von Darwin zur DNA. Heidelberg, Berlin, Oxford, S. 109-128
MINDEL, D. P. & MEYER, A. (2001) Homology evolving. Trends in ecology and evolution 16: 434-440.
MISHLER, BD et al. (1994) Phylogenetic relationships of the ‚Green Algae‘ and ‚Bryophytes‘. Ann. Miss. Bot. Gard. 81, S. 451-483
Neukamm, M. (2001): Die Evolutionstheorie und der moderne Antievolutionismus http://www.martin-neukamm.de/junker.html
NEUKAMM, M (2002) Die Ähnlichkeiten zwischen den Arten – Homologie als Evolutionsbeleg. Praxis der Naturwissenschaften – Biologie 7/51, S. 39-42
OSCHE, G (1979) Evolution. Grundlagen – Erkenntnisse, Entwicklungen der Abstammungslehre, Freiburg
PAGE, RD; COTTON, JA (2002): Vertebrate phylogenomics: reconciled trees and gene duplications. Pac Symp Biocomput. 2002, 536-547.
PROTHERO, D. (2017): Evolution – What the Fossils say and why it matters. Second edition. New York: Columbia University Press
REMANE, A. (1952): Die Grundlagen des natürlichen Systems, der vergleichenden Anatomie und der Phylogenetik. Leipzig: Geest & Portig K.-G.
REMANE, A et al. (1973) Evolution. Tatsachen und Probleme der Abstammungslehre. München
RIEDL, R (1990) Biologie der Erkenntnis. Berlin, Hamburg
RIEPPEL, O (1999) Einführung in die computergestützte Kladistik. München
SHUBIN N (1994) The phylogeny of development and the origin of homology. In: RIEPPEL und GRANDE (Hrsg.) (1994) Interpreting the hierarchy of nature. San Diego, S. 201-225
SUDHAUS, W und REHFELD, K (1992) Einführung in die Phylogenetik und Systematik. Stuttgart, Jena, New York
Talkorigins.org.: Index of Creationist Claims http://www.talkorigins.org/indexcc/list.html Kapitel Homologie: http://www.talkorigins.org/indexcc/CB/CB810.html
WÄGELE JW (2000) Grundlagen der Phylogenetischen Systematik. München
WELLS, J. (2000): Icons of Evolution, Washington DC: Regnery Publishing Inc
WIESER, W. (Hrsg. 1994): Die Evolution der Evolutionstheorie Von Darwin zur DNA, Spektrum Akademischer Verlag, Heidelberg, Berlin, Oxford
