
Text als pdf
Der Calvin-Zyklus ist das Herzstück der Photosynthese – der Prozess, durch den Pflanzen, Algen und viele Bakterien CO2 in Zucker verwandeln. Doch er ist weit mehr als nur Biochemie: Er ist das Ergebnis von Milliarden Jahren Evolution. Im Zentrum steht das Enzym RubisCO, eines der ältesten und häufigsten Proteine der Erde, das schon in frühen Cyanobakterien aktiv war. Trotz seiner entscheidenden Rolle ist RubisCO bemerkenswert ineffizient – ein Relikt aus einer Zeit, als die Erde noch kaum Sauerstoff kannte. Der Calvin-Zyklus selbst hat sich mehrfach angepasst und ist ein Paradebeispiel für die enge Verflechtung von Biochemie und Evolution. In diesem Video schauen wir uns an, wie dieser Zyklus entstand, warum er bis heute funktioniert – und warum er trotzdem alles andere als perfekt ist.
Die Lichtreaktion der Photosynthese wandelt die Lichtenergie in chemische Energie um. Diese wird dann bereitgestellt für die Fixierung des CO2 im Calvin-Zyklus, um Kohlenhydrate zu bilden. Der Calvin-Zyklus kann in Abwesenheit von Licht ablaufen, weswegen der auch als Dunkelreaktion oder lichtunabhängige Reaktion bezeichnet wird (Abb. 1). Der Zyklus wurde von den US-amerikanischen Biochemikern Melvin Calvin, Andrew A. Benson und James Alan Bassham in der Zeit von Ende der 1940er bis Mitte der 1950er Jahre entdeckt und entschlüsselt. Er ist die häufigste Form der CO2-Fixierung und macht etwa 99,5% der CO2-Fixierung aus. Er ist aber nicht der einzige. Die restlichen 0,5% teilen sich fünf weitere Kohlenstofffixierungs-Wege, von denen die meisten erdgeschichtlich deutlich älter sind als der Calvin-Zyklus (Raven 2009, Prywes et al. 2023, Bar-On & Milo 2019).
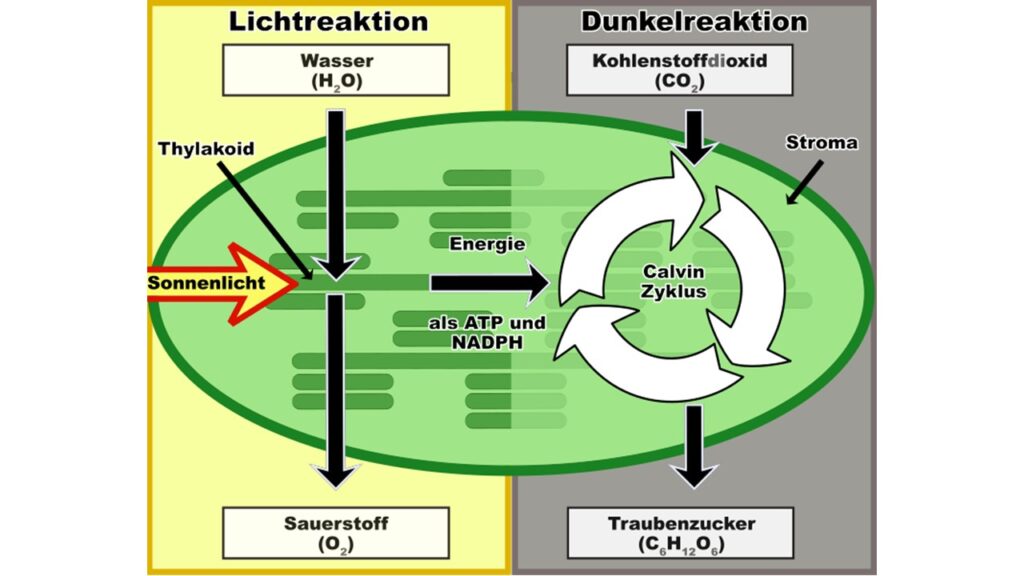
Abb. 1: Licht- und Dunkelreaktion
Die anderen Wege der CO2-Fixierung
Die wohl älteste Form der CO2-Fixierung haben wir in unserer Serie über den Ursprung des Lebens kennengelernt, den Acetyl-CoA-Weg (Abb. 2). Dessen Reaktion ist einfach und verläuft exergon. D. h. diesem Reaktionsweg muss keine zusätzliche Energie zugeführt werden, sondern bei ihm wird Energie frei. Die restlichen 5 CO2-Fixierungswege benötigen zusätzlich Energie, um CO2 zu fixieren; beim Calvin-Zyklus ist es z. B. die in der Photosynthese bereitgestellte Energie. Der Acetyl-CoA-Weg findet sich bei ursprünglichen Bakterien und Archaeen, wie den Acetogenen und Methanogenen (Berg et al. 2010, Fuchs 2011, Weiss et al. 2016).
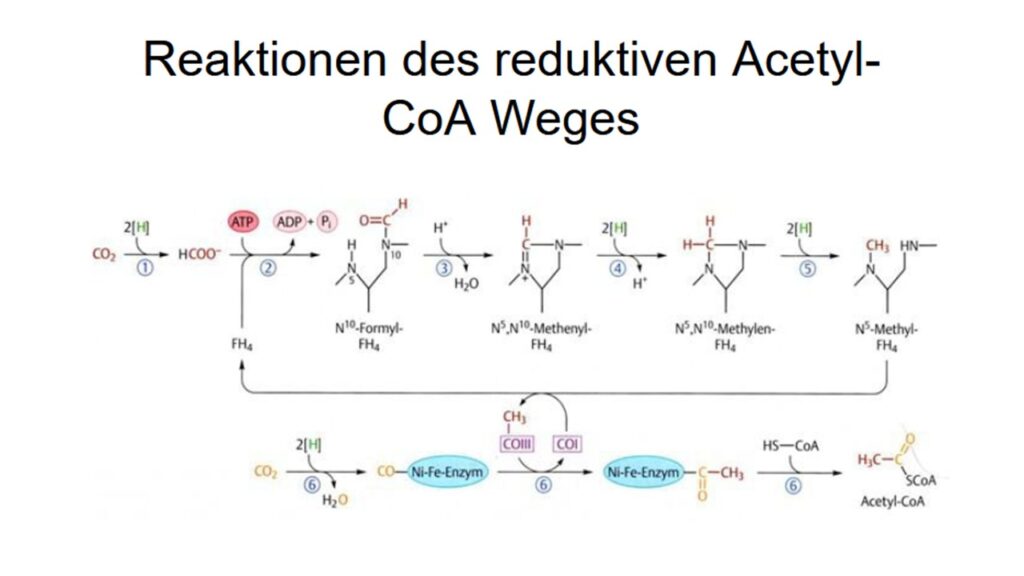
Abb. 2: reduktiver Acetyl-CoA-Weg
Eine andere Form der CO2-Fixierung, der reduktive bzw. rückwärtslaufende Krebszyklus, dürfte erdgeschichtlich ebenfalls sehr alt sein (Abb. 3). Er ist in Bezug zum Energieverbrauch und CO2-Fixierung sogar deutlich effizienter als die CO2-Fxierung der Photosynthese, kann aber seine Aufgabe – die Fixierung von CO2 – in Anwesenheit von Sauerstoff nicht durchführen und ist daher auf anaerobe Bedingungen beschränkt. Der rückwärtslaufende Krebszyklus ist quasi gesehen das Gegenstück des eigentlichen Krebszyklus der Zellatmung. In dieser Reaktionsrichtung spaltet er CO2 von organischen Molekülen ab, kann ein wenig Energie in Form von ATP speichern und außerdem Elektronen und Protonen für die Erzeugung von ATP in der Atmungskette bereitstellen (Fuchs 2011).
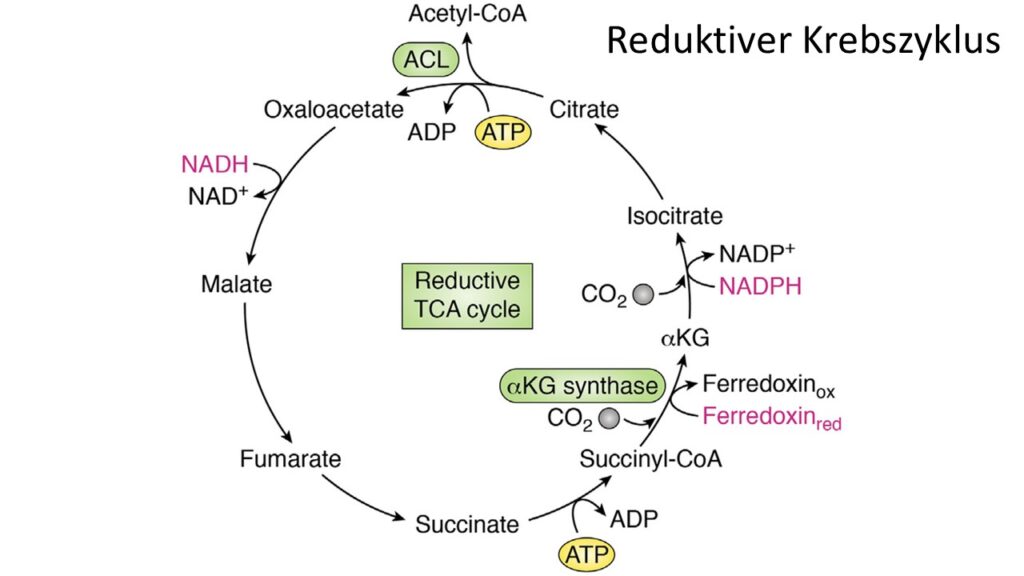
Abb. 3: reduktiver Krebszyklus
Drei weitere Formen der CO2 Fixierung – der 3-Hydroxypropionatzyklus, der 3-Hydroxypropionat/4-Hydroxybutyratzyklus und der Dicarboxylat/4-Hydroxybutyratzyklus sind nur auf einige wenige Bakterien oder Archaeen beschränkt und dürften den meisten Nicht-Biochemikern völlig unbekannt sein.
Der 3-Hydroxypropionatzyklus wurde erst in den 1980ern beim phototrophen Bakterium Chloroflexus aurantiacus nachgewiesen (Holo & Sirevag 1986, Strauss & Fuchs 1993, Fuchs 2011, Cai & Li 2016, Klatt et al. 2007, Ivanovsky et al. 1999; Abb. 4).
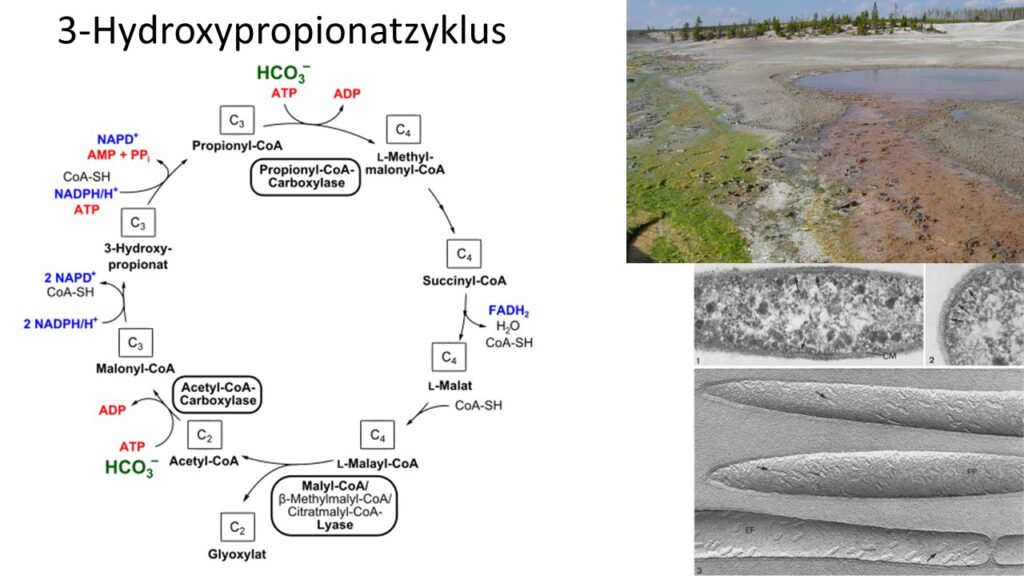
Abb. 4: 3-Hydroxypropionatzyklus
Beim 3-Hydroxypropionatzyklus werden zwei Moleküle CO2 in Form von Bicarbonat (HCO3−) zu einem Molekül Glyoxylat unter ATP- und NADPH-Verbrauch aufgebaut. Anschließend wird das entstandene Glyoxylat unter Fixierung eines weiteren Moleküls Bicarbonat zu Pyruvat umgesetzt. Ein Zwischenprodukt ist das namensgebende 3-Hydroxypropionat. Die Verwendung von Bicarbonat anstelle von CO2 kann damit erklärt werden, dass Chloroflexus aurantiacus in Gewässern mit leicht alkalischem Milieu wächst. Unter diesen Bedingungen ist die Konzentration an Bicarbonat höher als die von CO2. Eine Besonderheit des Zyklus ist, dass bi- und trifunktionale Enzyme beteiligt sind, d. h. einige Enzyme können zwei oder drei Reaktionen katalysieren. Der gesamte Zyklus erfolgt in 19 Reaktionen, jedoch sind nur 13 Enzyme an den Reaktionen beteiligt. Dieser Zyklus kann unter sauerstoffhaltigen (aeroben) stattfinden, da keine seiner involvierten Enzyme per se sauerstoffempfindlich sind. (Thauer 2007, Herter et al. 2002, Berg 2011, Zarzycki et al. 2009, Fuchs & Schlegel 2007, Madigan et al.2020).
Phylogenetischen Untersuchungen zur Folge ist diese Form der CO2-Fixierung mit 867 Mio. Jahren erdgeschichtlich jung und entwickelte sich erst nach der Freisetzung von Sauferstoff in die Atmosphäre. Wahrscheinlich wurde diese Fähigkeit durch horizontalen Gentransfer und der Umfunktionalisierung bestehender Enzyme verwirklicht (Shih et al. 2017). Die beiden anderen Formen der CO2-Fixierung sind eine Abwandlung des 3-Hydroxypropionatzyklus.
Der 3-Hydroxypropionat/4-Hydroxybutyratzyklus (Abb. 5) findet sich bei thermophilen Archeen der Gattung Metallosphaera, Acidianus und Sulfolobus (Berg et al. 2007). In vielen Reaktionsschritten ähnelt dieser Zyklus dem 3-Hydroxypropionatzyklus, es gibt aber auch einige Unterschiede, die u. a. auch zur Bildung des Zwischenprodukts 4-Hydroxybutyrat führen. Jedoch sind die darin beteiligten Enzyme bei beiden Zyklen phylogenetisch nicht verwandt und scheinen sich unabhängig voneinander entwickelt zu haben (Thauer 2007). Ein nur in diesem Zyklus involviertes Enzym, die 4-Hydroxybutyryl-CoA-Dehydratase, ist sensitiv gegenüber Sauerstoff, weshalb dieser Stoffwechselweg nur unter sauerstofffreien (anaeroben) Bedingungen ablaufen kann (Martins et al. 2004).
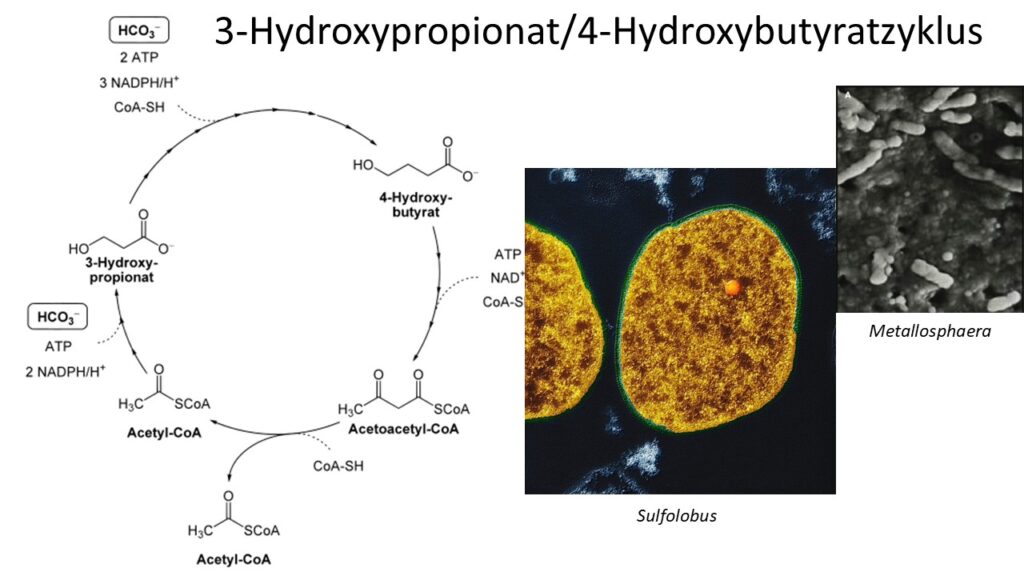
Abb. 5: 3-Hydroxypropionat/4-Hydroxybutyratzyklus
Der Kreislauf deckt sich auch teilweise mit dem kürzlich entdeckten Dicarboxylat/4-Hydroxybutyratzyklus (Abb. 6). Es wurde 2008 beim hyperthermophilen Archaeon Ignicoccus hospitalis entdeckt (Huber et al. 2008). Auch dieser läuft unter anaeroben Bedingungen ab.
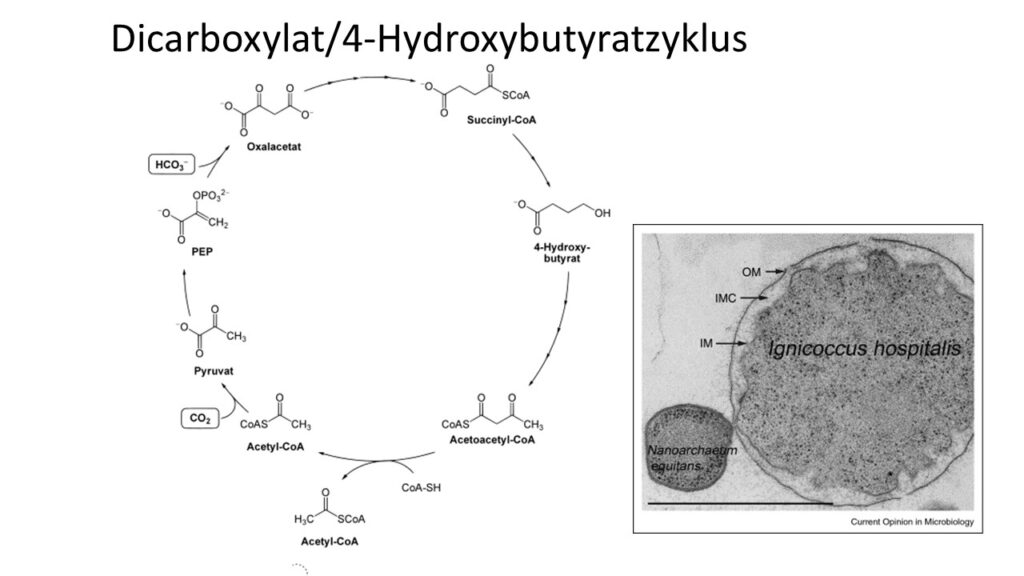
Abb. 6: Dicarboxylat/4-Hydroxybutyratzyklus
Calvin-Zyklus
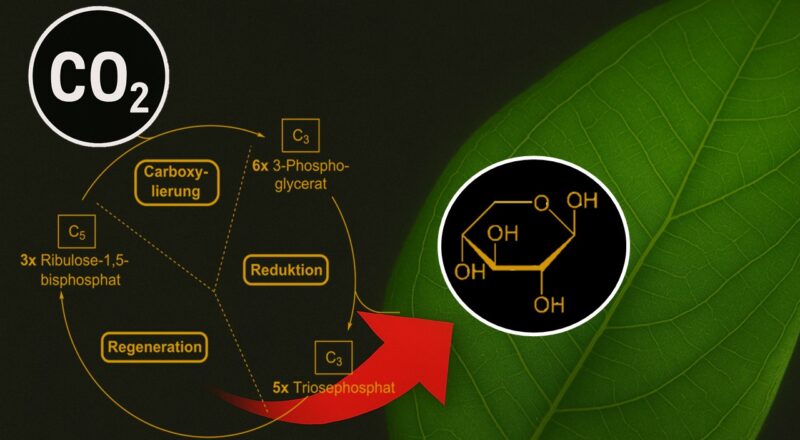
Der Calvin-Zyklus kann in drei Phasen untergliedert werden: Die CO2-Fixierung, die Reduktionsphase und die Regenerationsphase. CO2 aus der Luft diffundiert durch die Spaltöffnungen in die Blätter der Pflanze und von da in die Chloroplasten (Abb. 7, siehe: Campbell et al. 2016, Kadereit et al. 2021, Madigan et al. 2020, Munk 2008, Nelson & Cox 2010, Savada et al. 2019).
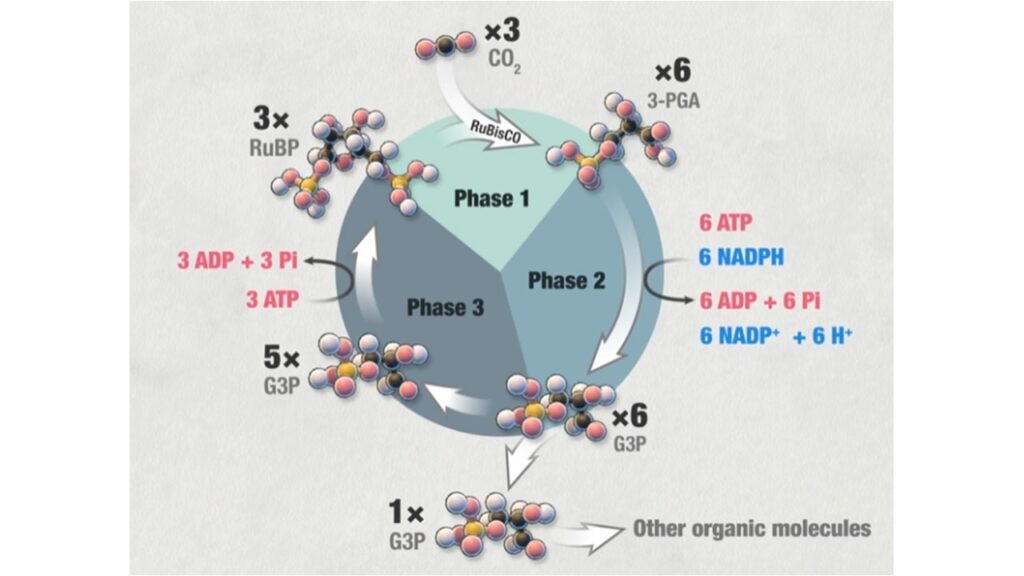
Abb. 7: Calvin-Zyklus
In der Fixierungsphase verbindet sich ein CO2-Molekül mit dem aus 5 Kohlenstoffatomen bestehenden Zucker Ribulose-1,5-bisphosphat (RubP). Diese Reaktion katalysiert ein Enzym mit dem Namen RuBisCO. Es entsteht ein instabiles Molekül mit 6 Kohlenstoffatomen, also ein C6-Körper, der in 2 Moleküle mit je 3 Kohlenstoffatomen, also C3-Körper, zerfällt, nämlich in 3-Phosphoglycerat.
Im nächsten Schritt – der Reduktionsphase – reagiert 3-Phosphoglycerat zu einer ebenfalls aus 3 Kohlenstoffatomen bestehenden Verbindung namens Glycerinaldehyd-3-phosphat (GAP). Diese Umwandlung benötigt die aus der Lichtreaktion der Photosynthese gewonnen Substrate ATP und NADPH, es ist also eine Reaktion die Energie benötigt.
Der letzte Reaktionsschritt – die Regenerationsphase – dient dazu Ribulose-1,5-bisphosphat wiederherzustellen. Dafür werden etwa 5/6 des gebildeten Glycerinaldehyd-3-phosphat verwendet. Auch hier ist wieder Energie in Form von ATP-Molekülen nötig. Jetzt kann der Zyklus wieder erneut durchlaufen werden, da Ribulose-1,5-bisphosphat für die CO2-Fixierung wieder zur Verfügung steht.
1/6 des gebildeten Glycerinaldehyd-3-phosphat aber verlässt den Zyklus und kann gemeinsam mit einem weiteren Molekül Glycerinaldehyd-3-phosphat zu einem aus 6 Kohlenstoffatomen bestehenden Zuckermolekül wie Glucose reagieren. Sie können dann in den Stoffwechsel eingeschleust und dort für die Energiegewinnung abgebaut werden. Unter günstigen Bedingungen, also wenn die Pflanzen mehr Glucose herstellt, als sie für ihren eigenen Energiebedarf benötigt, können die Zuckermoleküle als Baustoffe für verschiedene Makromoleküle wie Kohlenhydrate, Fette oder Proteine zur Verfügung gestellt werden. Die Kohlenhydrate können in den Chloroplasten in Stärkekörnern gespeichert und bei Bedarf mobilisiert werden. Für die Bildung eines Glucose-Moleküls, welches aus 6 Kohlenstoffatomen besteht, müssen 6 CO2 verbraucht werden, d. h. der Calvin-Zyklus muss 6 Mal durchlaufen werden, damit ein Glucose-Molekül gebildet werden kann. Insgesamt sind 18 Moleküle ATP und 12 NADPH notwendig. Es ist also ein Prozess sehr recht viel Energie kostet, mehr als z. B. beim reduktiven Krebszyklus, dort sind nur 2 Moleküle ATP und 8 NAD(P)H oder Ferredoxin für die CO2-Fixierung notwendig. Auch die anderen CO2-Fixierungen benötigen deutlich weniger ATP als der Calvin-Zyklus. Aber alle anderen CO2-Fixierungen laufen unter anaeroben Bedingungen ab bzw. sind in Form des 3-Hydroxpropionatzyklus erdgeschichtlich jünger, als der Calvin-Zyklus seinen Siegeszug in einer sauerstoffreichen Atmosphäre in der CO2-Fixierung feierte.
Evolution des Enzyms Rubisco
Das für den Calvn-Zyklus wichtigste Enzym ist Rubisco, welcher das CO2 mit den C5-Zucker Ribulose-1,5-bisphosphat verbindet. Rubisco steht für Ribulose-1,5-bisphosphat-Carboxylase-Oxygenase. Fast die Hälfte der Proteinmasse in den Blättern der Pflanzen besteht aus diesem Enzym (Feller et al. 2008), es dürfte damit das häufigste Protein der Welt sein. Rubisco ist so reichlich vorhanden, weil es für ein Enzym ungewöhnlich schlecht arbeitet. Normalerweise rasseln Enzyme mit unverständlicher Geschwindigkeit und katalysieren genau dieselbe Reaktion Hunderte oder Tausende von Malen pro Sekunde. Rubisco hingegen hat eine Umsatzrate von weniger als zehn Reaktionen pro Sekunde. Um die Photosyntheserate zu erhöhen, müssen die Pflanzen mehr Rubisco herstellen, weshalb es in großer Menge vorhanden ist. (Bar-Even et al 2011, Morell et al. 1992, Bathellier et al. 2018). Gleichzeitig muss jedoch berücksichtigt werden, dass „perfekte“ Enzyme einfachere Reaktionen katalysieren. CO2 ist ein kleines, hydrophobes, ungeladenes und träges Molekül, weswegen es eine Herausforderung ist, es als Substrat zu verwenden (Bathellier et al. 2018, Pryves et al. 2023).
Aber das ist noch nicht alles. Rubisco ist auch weniger spezifisch als die meisten anderen Enzyme: Es hat Schwierigkeiten, zwischen CO2 und Sauerstoff zu unterscheiden.
Der Name lässt schon vermuten, dass es eben nicht nur CO2 fixieren kann, sondern auch Sauerstoff, das ist die Oxygenase-Funktion von Rubisco. Und das kann für Pflanzen sehr problematisch werden. Fixiert RuBisCo Sauerstoff statt CO2 entsteht nicht 3-Phosphoglycerat, sondern das C2-Molekül 2-Phosphoglycolat. Auch wenn der Name ähnlich klingt, ist 2-Phosphoglycolat für die Pflanze unbrauchbar. Dieses muss in einem energieaufwendigen Prozess wieder in 3-Phosphoglycerat umgewandelt werden. Dieser energiezehrende Prozess ist als Lichtatmung oder Photorespiration bekannt (Abb. 8). Bei heißem und trockenem Klima beginnen sich die Poren, die den Gasaustausch in den Blättern ermöglichen, zu schließen, wodurch der Wasserverlust eingeschränkt wird. Dies stellt Rubisco vor ein ernsthaftes Problem, da der Sauerstoff im Blattinneren eingeschlossen ist, während der CO2-Gehalt sinkt. Die Pflanzen können unter diesen Bedingungen ein Viertel ihres Ertrags einbüßen, da sie mehr Sauerstoff fixieren. Unter Umständen kann die Pflanze so mehr Energie verbrauchen als sie Glucose gewinnen kann und absterben. Entsprechend haben einige Pflanzengruppen interessante Strategien entwickelt diese Lichtatmung zu minimieren – dazu später mehr. Denn wir müssen uns fragen, wie ein solch schlecht arbeitendes Enzym, dass unter ungünstigen Bedingungen den sicheren Tod der Pflanze einleitet, überhaupt für die dominante Fixierung von CO2 genutzt werden konnte. Betrachten wir zuerst die Struktur von Rubisco, denn es gibt verschiedene Typen.
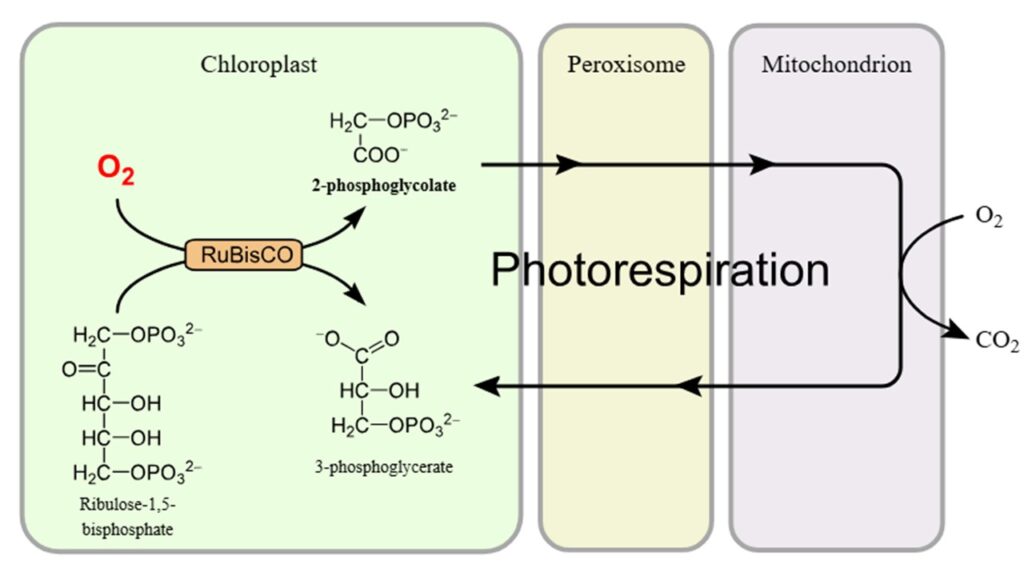
Abb. 8: Lichtatmung
Typ1-Rubisco kommt bei Pflanzen, Cyanobakterien und einigen anderen Bakterien vor. Dieses besteht aus zwei Proteinuntereinheiten, der großen L-Kette und der kleinen S-Kette. Das Gen für die L-Kette findet sich im Chloroplasten, die S-Kette im Kerngenom der Pflanze, deren Genprodukte aus dem Zellplasma in die Chloroplasten transportiert werden (Vitilin Gruber & Feiz 2018, Chase et al. 1993, Dhingra et al. 2004, Yoon et al. 2001). Die enzymatisch aktiven Bindungsstellen befinden sich in den großen Ketten und dort finden sich Magnesium-Ionen, die die Bindung mit CO2 ermöglicht (Lodish et al. 2000, Stec 2012).
Rubisco des Typs 2 kommt bei einigen Proteobakterien, Archaeen und Dinoflagellaten (Anderson et al. 1968) vor. Sie hat keine kleinen Untereinheiten und ist weniger spezifisch für die Bindung von Sauerstoff.
Rubiscos des Typs 3 wurden 1999 entdeckt (Watson et al. 1999, Ezaki et al. 1999); sie unterscheiden sich von den anderen Formen durch ihr ausschließliches Vorkommen in Archaeen und sind nicht am Calvin-Zyklus beteiligt. Auch andere Organismen verwenden ihre Rubsico-Enzyme nicht nur im Calvin-Zyklus (Liu et al. 2017). Einige Pflanzen verwenden Rubisco, um die Effizienz der Biosynthese von Ölen im Samen zu verbessern (Schwender et al. 2004). In einigen heterotrophen Bakterien kann der Calvin-Zyklus als sekundäre Elektronensenke dienen (Liu et al. 2017). Rubiscos des Typs 2 und 3 spielen typischerweise eine Rolle bei Nukleosid-Stoffwechslwegen (Wringhton et al. 2016, Sato et al. 2007) oder dem reduktiven Hexulose-Phosphat-Weg (Kono et al. 2017). Sie werden in Genomen gefunden, denen offensichtlich Phosphoribulokinase (prk)-Gene fehlen, die an der Regulation von Rubisco im Calvinzyklus beteiligt sind.
Im Jahr 2000 wurden Rubiscos des Typ 4 entdeckt (Hanson et al. 2001) und werden auch als Rubisco-ähnliche Proteine bezeichnet, da sie andere Reaktionen katalysieren. Aufgrund ihrer eindeutigen Homologie zu den anderen Rubiscos waren sie höchstwahrscheinlich evolutionäre Vorläufer (Hanson et al. 2001). Bei verschiedenen Kladen, die über Rubisco-ähnliche Proteine verfügen wurden eine Reihe unterschiedlicher chemischer Reaktionen identifiziert, die sie katalysieren (Kim et al. 2018, Carter et al. 2018, Kreel & Tabita 2007, Dey et al. 2015, Forlov et al. 2019). Trotz extremer Unterschiede in der Sequenz sind die zugrundeliegenden Mechanismen der verschiedenen Rubisco-ähnlichen Proteine und Rubisco ähnlich genug, um die Reaktionen der jeweils anderen zu katalysieren.
Phylogenetische Sequenzen haben den evolutionären Weg von Rubicso enträtselt. So zeigen viele Analysen, dass Rubisco vom Typ 3 tief im evolutionären Stammbaum bei den Archeen verwurzelt ist, was zur Hypothese führt, dass die Evolution von Rubisco hier begann und über viele Fälle von horizontalem Gentransfer und strukturellen Innovationen zu der heute beobachteten phylogenetischen Verteilung führte (Tabita et al. 2008). Nach dieser Hypothese war der Vorfahre ein echter Rubisco. Einer Hypothese zur Folge entwickelte sich Rubisco aus einem Methionin-Salvage-Enzym (Ashida et al. 2008) vor mehr als 2,9 Milliarden Jahren (Nisbet et al. 2007), also vor dem großen Oxygenierungsereignis (Warcke et al. 2020). Der Methionin-Salvage-Stoffwechselweg ist eine Abfolge von sechs chemischen Reaktionen, die das Ausgangsprodukt 5′-Methylthioadenosin (MTA) in das Endprodukt L-Methionin umwandeln. Er ist zumindest in den auf Methionin angewiesenen Organismen lebenswichtig zur Rückgewinnung des Schwefelatoms, dessen Assimilation energieaufwändig ist (Wray & Abeles 1995, Pirkov et al. 2008, Miyazaki & Yang 1987).
Eine alternative Hypothese, besagt, dass sich Rubisco aus einem Rubisco-ähnlichen Protein, also Typ IV, entwickelt hat, welches heterotrophe Stoffwechselwege katalysierte und die Fahrigkeit CO2 zu fixieren später evolvierte (Erb & Zarzycki 2018). Argumentiert wird hierbei, dass die ältesten Formen von Rubisco, jene von Typ 3, nur bei den stammesgeschichtlich ursprünglichen Archeen vorkommen und nicht am Calvin-Zyklus beteiligt sind, sondern die Assimilation von Ribonukleosiden katalysieren (Aono et al. 2012, 2015, Sato et al. 2007, Abb. 9). Im so genannten Pentosebisphosphatweg in Archaeen werden die Riboseeinheiten von Adenosin und anderen Nukleosiden mittels Rubisco über Ribulose-1,5-bisphosphat zu zwei Molekülen 3-Phosophglycerat verstoffwechselt. Die Assimilation von Ribose über einen Weg, der eine Carboxylierungsreaktion beinhaltet, erhöht die Biomasseerträge aufgrund der zusätzlichen Fixierung von anorganischem Kohlenstoff (d. h. CO2). Die Entstehung der Urform III von RubisCO und seine Einbindung in den Pentosebisphosphatweg für die Nukleotidassimilation wäre also in der alten Biosphäre von direktem Vorteil gewesen.
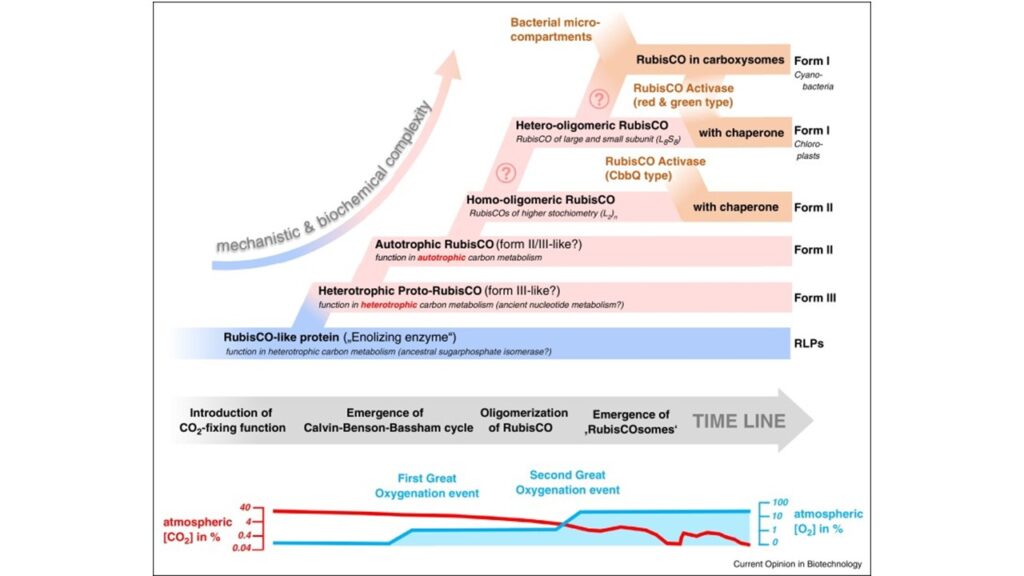
Abb. 9: Evolution der Rubiscos
Unterstützt wird diese Hypothese, dass es eine Reihe von Bakterien gibt, die in anaeroben Umgebungen häufig anzutreffen sind und dort organische Kohlenstoffverbindung mittels Gärungen verstoffwechseln. Mehrere dieser Bakterien verfügen über einen Nukleotid-Stoffwechsel auf Rubisco-Basis, der dem Pentosebiphosphatweg der Archeen ähnelt. Wahrscheinlich wurden die Gene für die Rubisco-Enzyme mittels horizontalem Gentransfer übertragen (Wrighton et al. 2017).
Es ist gut dokumentiert, dass horizontaler Gentransfer bei verschiedenen Rubiscos weit verbreitet ist (Tabita et al. 2007, Jaffe et al. 2019, Delwiche & Palmer 1996, Paoli et al. 1998). Pflanzen erwarben ihre Rubiscos Cyanobakterien während der Endosymbiose, bei dem die kleine Untereinheit ins Kerngenom wanderte (Bouvier et al. 2022). Proteobakterien haben oft mehrere Rubiscos der Form I und II in ihren Genomen, mit unterschiedlichen biochemischen Eigenschaften (Badger & Bek 2008). Bei Nicht-Grünen Algen fanden weitere Formen des horizontalen Gentransfers statt (Delwiche & Palmer 1996, Whitney et al. 1995). Schließlich wurde kürzlich eine Gruppe von Rubiscos in den Genomen von Bakteriophagen (Myoviridae-Phagen) und ihren Wirten, den Beckwithbakterien, gefunden (Jaffe et al. 2019). Dies legt die Vermutung nahe, dass Viren als Vehikel für den horizontalen Gentransfer zwischen den Lebensbereichen dienen.
Wie schnell könnte sich eine erste Version des Calvin-Zyklus entwickelt haben? Erstaunlich schnell! Jüngste Experimente zeigen, dass sich ein funktionsfähiger Calvin-Zyklus bemerkenswert schnell und reproduzierbar entwickeln kann, wenn RubisCO zusammen mit Phosphoribulokinase in Escherichia coli heterolog exprimiert wird und Selektionsdruck ausgeübt wird. Es braucht nur hundert Generationen und eine Handvoll Mutationen, um den heterotrophen Organismus E. coli in einen halbautotrophen Organismus zu verwandeln, in dem alle Zucker nur aus CO2 gebildet werden (Antonovsky et al. 2016, Gleizer et al. 2019, Herz et al. 2017).
Lichtatmung und Evolution der C4 und CAM-Photosynthese
Als Rubisco entstand war der CO2 -Gehalt in der Atmosphäre viel höher als heute, während die Sauerstoffkonzentration nur einen Bruchteil der heutigen 21 % betrug. Das bedeutet, dass die schlechte Unterscheidungsfähigkeit von Rubisco damals kein Problem war; es war viel wahrscheinlicher, dass sie CO2 als Sauerstoff bindet. Heute ist die Situation umgekehrt, mit Sauerstoff -Konzentrationen, die Hunderte Male höher sind als CO2, vor allem in den Blättern, wo CO2 aufgenommen und Sauerstoff als Abfallprodukt ausgeschieden wird (Fischer et al. 2016). Das führt zum oben beschriebenen Problem der Lichtatmung. Interessant ist jedoch, dass eine Reihe von Studien belegen, dass die Lichtatmung für die Stickstoffaufnahme notwendig sein kann (Busch et al. 2018, Bloom & Lancaster 2018) und Stickstoff ist oft der begrenzende Faktor für das Wachstum (Sinclair et al. 2019).
Die Notwendigkeit die Reaktion von Ferredoxin mit Sauerstoff zu verhindern, könnte die Neigung von Rubisco erklären, Sauerstoff durch den scheinbar sinnlosen Prozess der Lichtatmung zu binden. Ferredoxin ist ein eisenhaltiges Protein und Bestandteil der Elektronentransportkette der Lichtreaktion. Seine Aufgabe besteht darin der letzte Elektronenakzeptor der Elektronentransportkette zu sein und NADP+ zu NADPH/H+ zu reduzieren. Stellen wir uns nun vor, was im Fall von Rubisco passiert, wenn der CO2-Gehalt sinkt, während der Sauerstoffgehalt im Blattinnern steigt. Rubisco muss seine Arbeit verlangsamen, weil sein Substrat, das CO2 knapp wird. Das bedeutet aber auch, dass das NADPH seine Elektronen nicht weitergeben kann um NADP+ zu regenerieren. Infolgedessen kann auch Ferredoxin seine Elektronen nicht mehr abgeben und wird mit Sauerstoff reaktiv, und zwar genau dann, wenn der Sauerstoffgehalt steigt. Um die Katastrophe abzuwenden, verbraucht Rubisco stattdessen Sauerstoff. Die Lichtatmung wandelt NADPH wieder in NADP+ um, so dass Ferredoxin seine Elektronen wieder abgeben kann. Es könnte also sein, dass die Lichtatmung wie ein Sicherheitsventil wirkt, indem sie den Gehalt an reaktivem Ferredoxin und Sauerstoff gleichzeitig senkt und so eine drohende Katastrophe abwendet. Sicherlich kann dies die Pflanzen einen beträchtlichen Ertrag kosten, aber zumindest überleben sie, um an einem anderen Tag zu sterben. Die Strafe für das Ersetzen von Rubisco durch ein „verbessertes“ Enzym könnte der plötzliche Tod sein.
Nichtsdestotrotz – ein zu viel an Lichtatmung kann für Pflanzen ein Problem darstellen und einige Pflanzen haben Strategien entwickelt, die Kosten der Lichtatmung zu minimieren – besonders in trockenen und heißen Standorten. Dies geschieht mittels C4 und CAM-Photosynthese (Abb. 10). Ziel ist RuBisCO immer einer hohen Konzentration an CO2 auszusetzen, damit es nicht die Möglichkeit hat, an Sauerstoff zu binden. Die spezielle Anpassung der C4– Pflanzen ist eine räumliche Trennung zwischen der CO2-Fixierung und dem eigentlichen Calvin-Zyklus.
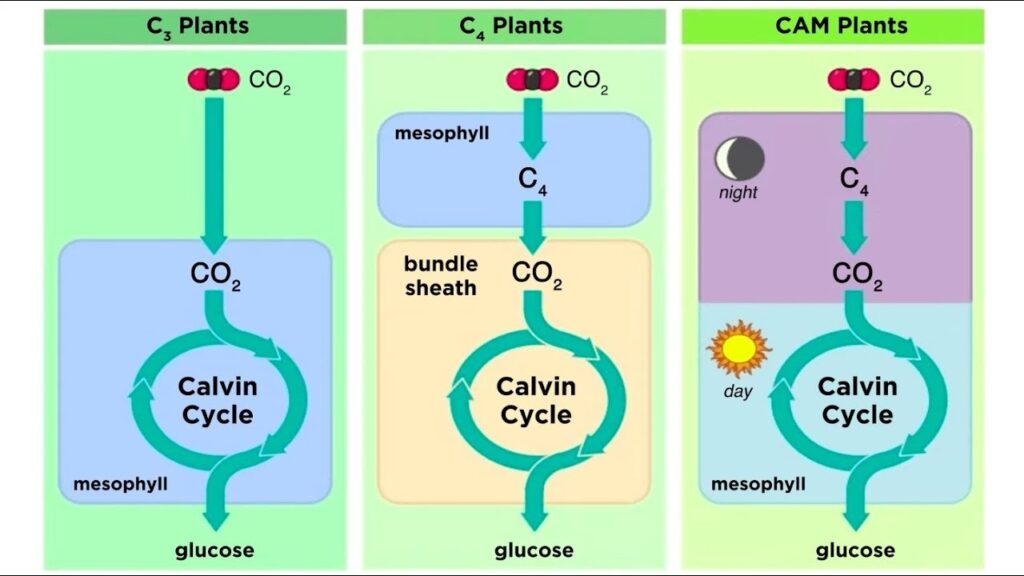
Abb. 10: C3, C4 und CAM-Photosynthese
In den Mesophyllzellen findet hierfür eine Vorfixierung des CO2 statt, wobei dieses mit Phosphoenolpyruvat (PEP), einem Stoffwechselzwischenprodukt der Glykoloyse, fixiert und den C4-Zucker Oxalacetat bindet, der anschließend in Malat oder Asparat umgewandelt wird. Da bei dieser Vorfixierung ein Zucker mit vier Kohlenstoffatomen entsteht, spricht man von C4-Photosynthese.
Die C4-Zucker werden dann in die Bündelscheidenzellen transportiert, die sich wie ein Kranz um die Mesophyllzellen anordnen. Hier wird CO2 vom C4-Körper abgespalten und der Calvin-Zyklus kann wie gewohnt stattfinden.
Bei CAM-Pflanzen findet ebenfalls eine Vorfixierung des CO2 statt, bei der ein C4-Körper entsteht. Im Gegensatz zu C4-Pflanzen findet hier aber keine räumliche, sondern zeitliche Trennung zwischen CO2-Vorfixierung und Calvin-Zyklus statt.
Nachts öffnen die CAM-Pflanzen ihre Spaltöffnungen, damit CO2 in die Blätter diffundieren kann. Dort wird dann das CO2 ähnlich wie bei C4-Pflanzen vorfixiert. Die entstehende C4-Säure wird in die Vakuolen der Zellen transportiert und dort gespeichert. Tagsüber schließen die Pflanzen wieder ihre Spaltöffnungen, um den Wasserverlust zu verringern. Die C4-Säure kann nun aus den Vakuolen transportiert und CO2 abgespalten werden. Dadurch kann der Calvin-Zyklus wieder wie gewohnt ablaufen.
Etwa 3% der Arten der Bedecktsamer betreiben eine C4-Photosynthese und machen etwa 5% der Biomasse aus. Sie fixieren aber 23% des CO2 (Bond et al. 2005, Kellogg 2013). Hierzu gehören vornehmlich Gräser, die aus wärmeren Gebieten mit längeren Trockenperioden stammen, z. B. Mais, Zuckerrohr oder Hirse. 46 % der Gräser sind C4-Arten und machen zusammen 61 % der C4-Arten aus. C4 ist in der Familie der Gräser über zwanzig Mal unabhängig voneinander in verschiedenen Unterfamilien, Stämmen und Gattungen aufgetreten (Grass Phylogeny Working Group II 2012). C4-Photosynthese kommt aber auch in anderen Pflanzenfamilien vor und ist wahrscheinlich 61-mal unabhängig voneinander in 19 Pflanzenfamilien entstanden (Sage 2004).
Bei Nadelbäumen, Farnen und Moosen kommt eine C4-Photosynthese gar nicht vor, bei Laubbäumen nur in geringen Ausnahmefällen. Diese Pflanzen nutzen die C3-Photosynthese, also die Photosynthese ohne eine Vorfixierung des CO2 (Sage 2017).
Gegenüber C3-Pflanzen zeichnen sich C4-Pflanzen bei Wasserknappheit, hohen Temperaturen und Sonneneinstrahlung aus. C4-Pflanzen dominieren also eher trockene Gebiete, z. B. die Savannen Afrikas oder Australiens (Sage & Monson 1999). In tropischen Regenwäldern dominieren hingegen die C3-Pflanzen, was nicht verwundern sollte, da die C4-Photosynthese in Bäumen praktisch gar nicht vorkommt.
C4-Pflanzen entstanden vor etwa 35 Millionen Jahren (Sage 2004) im Oligozän (und erlangten erst vor etwa 6 bis 7 Millionen Jahren, im Miozän, ökologische Bedeutung (Osborne & Beerling 2006). In diesem Zeitraum wurde das Klima trockener und die Savannen breiteten sich in weiten Regionen der heutigen Tropen aus, wo die hohe Sonneneinstrahlung einen Vorteil gegenüber dem C3-Stoffwechsel bot (Edwards & Smith 2010, Osborne & Freckelton 2009).
Zu den CAM-Pflanzen gehören oft sukkulente Arten, also Pflanzen, deren Blätter oder anderen Organe viel Wasser speichern und an trockene Standorte angepasst sind. Hierzu zählen Vertreter der Dickblattgewächse (Crassulaceae), nach denen dieser Typ der CO2-Fixierung benannt ist, sondern auch viele Arten aus den Familien Cactaceae, Agavaceae und Euphorbiaceae. CAM-Pflanzen kommen vorwiegend in Trockengebieten vor, sind aber auch in tropischen Regenwäldern zu finden. Denn viele Aufsitzerpflanzen, sog. Epiphyten, also Pflanzen, die auf anderen Pflanzen wachsen, leiden auch unter Wassermangel und hohen Temperaturen. Hierzu zählen z. B. viele Orchideen und Bromelien. Entsprechend kommt auch bei ihnen die CAM-Photosynthese relativ häufig vor.
Wenn die C4- und CAM-Photosynthese mehrmals unabhängig voneinander in verschiedenen Pflanzengruppen entstanden ist, stellt sich die Frage, welche evolutionären Vorbedingungen eine solche Entwicklung begünstigen. Überraschenderweise zeigen eine Reihe von Studien, dass Veränderungen der Anatomie und Zellbiologie der C4-Photosynthese vorausgingen, die nicht im direkten Zusammenhang mit dieser stehen (Williams et al. 2013, Christin et al 2012, Sage et al. 2018). So nimmt die Möglichkeit eine C4-Photosynthese zu entwickeln zu, wenn das Leitbündelscheidengewebe besonders gut ausgeprägt ist. Leitbündel sind für den Transport von Wasser, Mineralsalzen und organischen Substanzen innerhalb einer Pflanze zuständig. Das Grund- und Festigungsgewebe um die Leitbündel bezeichnet man als Leitbündelscheide. Eine Reihe von Enzymen, die an der Vorfixierung des CO2 bei C4-Pflanzen beteiligt sind, wie Phosphoenolpyruvat-Carboxylase und Malat-Dehydrogenase, erfüllen bei C3-Pflanzen andere Aufgaben (Aubry et al. 2011), so dass die frühen Veränderungen in der Häufigkeit und Lokalisierung dieser Enzyme in C4-Linien möglicherweise durch nichtphotosynthetische Einflüsse verursacht wurden.
Die C4-Photosynthese ist ein hervorragendes Beispiel dafür, wie unabhängige Abstammungslinien mit einem breiten Spektrum von Vorläuferphänotypen zu ähnlichen komplexen Merkmalen konvergieren können. Mehrere Studien zu einfacheren Merkmalen haben gezeigt, dass die Konvergenz zu einem Phänotyp durch verschiedene Genotypen und somit durch nicht homologe molekulare Mechanismen in unabhängigen Abstammungslinien spezifiziert werden kann.
Die Photosynthese ermöglichte nicht nur die gewaltige Leistung mittels Lichtenergie Wasser zu spalten, um so Energie für die Fixierung von CO2 zu gewinnen. Ihr Abfallprodukt, Sauerstoff, ermöglicht auch unser Leben. Doch zu Beginn war Sauerstoff ein tödliches Gift, welches das erste Massenaussterben der Erdgeschichte einleitete. Nur wenige konnten den Sauerstoff für seine Zwecke zur Energiegewinnung nutzen. Aber wie? Das erfahren wir in der letzten Episode unserer Geschichte zur Evolution der Photosynthese.
Literatur
Anderson LE, Price GB, Fuller RC. (1968). Molecular Diversity of the Ribulose-1,5-Diphosphate Carboxylase from Photosynthetic Microorganisms
Antonovsky N, Gleizer S, Noor E, Zohar Y, Herz E, Barenholz U, et al. (2016). Sugar Synthesis from CO2 in Escherichia coli. Cell 166: 115–125.
Aono R, Sato T, Imanaka T, Atomi H (2015). A pentose bisphosphate pathway for nucleoside degradation in Archaea. Nat Chem Biol, 11: 355-360
Aono R, Sato T, Yano A, Yoshida S, Nishitani Y, Miki K, Imanaka T, Atomi H (2012). Enzymatic characterization of AMP phosphorylase and ribose-1,5-bisphosphate isomerase functioning in an archaeal AMP metabolic pathway. J Bacteriol, 194: 6847-6855
Ashida H, Saito Y, Nakano T, Tandeau de Marsac N, Sekowska A, Danchin A, et al. (2008). RuBisCO-like proteins as the enolase enzyme in the methionine salvage pathway: functional and evolutionary relationships between RuBisCO-like proteins and photosynthetic RuBisCO. J Exp Bot. 59: 1543–1554.
Aubry S Brown NJ Hibberd JM (2011). The role of proteins in C3 plants prior to their recruitment into the C4 pathway J Exp Bot 62:3049–3059.
Badger MR, Bek EJ. (2008). Multiple Rubisco forms in proteobacteria: their functional significance in relation to CO2 acquisition by the CBB cycle. J. Exp. Bot. 59(7):1525–41
Bar-Even A, Noor E, Savir Y, Liebermeister W, Davidi D, et al. (2011). The moderately efficient enzyme: evolutionary and physicochemical trends shaping enzyme parameters. Biochemistry. 50(21):4402–10
Bar-On YM, Milo R (2019). The global mass and average rate of rubisco. Proc. Natl. Acad. Sci. U. S. A.
Bathellier C, Tcherkez G, Lorimer GH, Farquhar GD. (2018). Rubisco is not really so bad. Plant Cell Environ. 41(4):705–16
Berg IA (2011). Ecological aspects of the distribution of different autotrophic CO2 fixation pathways. Applied and Environmental Microbiology. 77 (6): 1925–36.
Berg IA, Kockelkorn D, Buckel W, Fuchs G (2007). A 3-hydroxypropionate/4-hydroxybutyrate autotrophic carbon dioxide assimilation pathway in Archaea. Science. 318 (5857): 1782–6.
Berg, I. A., Kockelhorn, D., Ramos-Vera, W. H., Say, R. F., Zarzyncki, J., Hügler, M., et al. (2010). Autotrophic carbon fixation in archaea. Nat. Rev. Microbiol. 8, 447–460.
Bloom AJ, Lancaster KM. (2018). Manganese binding to Rubisco could drive a photorespiratory pathway that increases the energy efficiency of photosynthesis. Nat Plants. 4(7):414–22
Bond, J., Woodward, F. I., Midgley, G. F. (2005): The global distribution of ecosystems in a world without fire. New Phytologist. 165(2): 525–538.
Bouvier JW, Emms DM, Kelly S. (2022). Slow molecular evolution of rubisco limits adaptive improvement of CO2 assimilation. Work. Pap.
Busch FA, Sage RF, Farquhar GD. (2018). Plants increase CO2 uptake by assimilating nitrogen via the photorespiratory pathway. Nat Plants. 4(1):46–54
Cai, Z & Li, Y (2016). Synthetic biology for CO2 fixation. Science China. Life sciences. 59. 10.1007/s11427-016-0304-2.
Campbell, N. A. et al. (2016): Biologie. München: Pearson.
Carter MS, Zhang X, Huang H, Bouvier JT, Francisco BS, et al. (2018). Functional assignment of multiple catabolic pathways for D-apiose. Nat. Chem. Biol. 14(7):696–705
Chase MW, Soltis DE, Olmstead RG, Morgan D, Les DH, Mishler BD, et al. (1993). Phylogenetics of Seed Plants: An Analysis of Nucleotide Sequences from the Plastid Gene rbcL. Annals of the Missouri Botanical Garden. 80 (3): 528–580.
Christin, P.-A.; Osborne, C. P.; Chatelet, D. S.; Columbus, J. T.; Besnard, G.; Hodkinson, T. R.; Garrison, L. M.; Vorontsova, M. S.; Edwards, E. J. (2012). Anatomical enablers and the evolution of C4 photosynthesis in grasses. Proceedings of the National Academy of Sciences. 110 (4): 1381–1386
Delwiche CF, Palmer JD. (1996). Rampant horizontal transfer and duplication of rubisco genes in eubacteria and plastids. Mol. Biol. Evol. 13(6):873–82
Dey S, North JA, Sriram J, Evans BS, Tabita FR. (2015). In Vivo Studies in Rhodospirillum rubrum Indicate That Ribulose-1, 5-bisphosphate Carboxylase/Oxygenase (Rubisco) Catalyzes Two Obligatorily Required and Physiologically Significant Reactions for Distinct Carbon and Sulfur Metabolic Pathways. J. Biol. Chem. 290(52):30658–68
Dhingra A, Portis AR, Daniell H (2004). Enhanced translation of a chloroplast-expressed RbcS gene restores small subunit levels and photosynthesis in nuclear RbcS antisense plants. Proceedings of the National Academy of Sciences of the United States of America. 101 (16): 6315–6320.
Edwards EJ, Smith SA (2010). Phylogenetic analyses reveal the shady history of C4 grasses. Proceedings of the National Academy of Sciences of the United States of America. 107 (6): 2532–7.
Erb TJ, Zarzycki J. (2018). A short history of RubisCO: the rise and fall (?) of Nature’s predominant CO2 fixing enzyme. Curr. Opin. Biotechnol. 49:100–107
Ezaki S, Maeda N, Kishimoto T, Atomi H, Imanaka T. (1999). Presence of a structurally novel type ribulose-bisphosphate carboxylase/oxygenase in the hyperthermophilic archaeon, Pyrococcus kodakaraensis KOD1. J. Biol. Chem. 274(8):5078–82
Feller U, Anders I, Mae T (2008). Rubiscolytics: fate of Rubisco after its enzymatic function in a cell is terminated. Journal of Experimental Botany. 59 (7): 1615–1624.
Fischer WW, Hemp J, Johnson JE. (2016). Evolution of Oxygenic Photosynthesis. Annu. Rev. Earth Planet. Sci. 44(1):647–83
Frolov EN, Kublanov IV, Toshchakov SV, Lunev EA, Pimenov NV, et al. (2019). Form III RubisCO- mediated transaldolase variant of the Calvin cycle in a chemolithoautotrophic bacterium. Proc. Natl. Acad. Sci. U. S. A., p. 201904225
Fuchs, G. (2011): Alternative pathways of carbon dioxide fixation: insights into the early evolution of life? Annu. Rev. Microbiol. 65, 631–658.
Fuchs G, Schlegel HG (2007): Allgemeine Mikrobiologie. Thieme Verlag Stuttgart; 8. Auflage
Gleizer S, Ben-Nissan R, Bar-On YM, Antonovsky N, Noor E, Zohar Y, et al (2019). Conversion of Escherichia coli to Generate All Biomass Carbon from CO2. Cell 179: 1255–1263.e12.
Grass Phylogeny Working Group II (2012). New grass phylogeny resolves deep evolutionary relationships and discovers C4 origins. The New Phytologist. 193 (2): 304–12.
Hanson TE, Tabita FR. (2001). A ribulose-1,5-bisphosphate carboxylase/oxygenase (RubisCO)-like protein from Chlorobium tepidum that is involved with sulfur metabolism and the response to oxidative stress. Proc. Natl. Acad. Sci. U. S. A. 98(8):4397–4402
Herter S, Busch A, Fuchs G (2002). L-Malyl-coenzyme A lyase/beta-methylmalyl-coenzyme A lyase from Chloroflexus aurantiacus, a bifunctional enzyme involved in autotrophic CO2 fixation. Journal of Bacteriology. 184 (21): 5999–6006.
Herz E, Antonovsky N, Bar-On Y, Davidi D, Gleizer S, Prywes N, et al. (2017). The genetic basis for the adaptation of E. coli to sugar synthesis from CO 2. Nat Commun 8: 1705.
Holo H, Sirevåg R (1986): Autotrophic growth and CO2 fixation of Chloroflexus aurantiacus. Arch. Microbiol. 145(2), S. 173–180
Huber H, Gallenberger M, Jahn U, Eylert E, Berg IA, Kockelkorn D, et al. (2008). A dicarboxylate/4-hydroxybutyrate autotrophic carbon assimilation cycle in the hyperthermophilic Archaeum Ignicoccus hospitalis. Proceedings of the National Academy of Sciences of the United States of America. 105 (22): 7851–6.
Ivanovsky RN et al. (1999): Evidence for the presence of the reductive pentose phosphate cycle in a filamentous anoxygenic photosynthetic bacterium, Oscillochloris trichoides strain DG-6. In: Microbiology 145 (7), 1743–1748
Jaffe AL, Castelle CJ, Dupont CL, Banfield JF. (2019). Lateral Gene Transfer Shapes the Distribution of RuBisCO among Candidate Phyla Radiation Bacteria and DPANN Archaea. Mol. Biol. Evol. 36(3):435–46
Kadereit, J. W.; et al. (2021): Strasburger − Lehrbuch der Pflanzenwissenschaften. Berlin Heidelberg New York: Springer-Verlag, 38. Auflage.
Kellogg, E. A. (2013): C4 photosynthesis. Current Biology. 23(14): R594–R599.
Klatt CG et al. (2007): Comparative genomics provides evidence for the 3-hydroxypropionate autotrophic pathway in filamentous anoxygenic phototrophic bacteria and in hot spring microbial mats. In: Environ Microbiol. 9(8), 2067–2078
Kim SM, Lim HS, Lee SB. (2018). Discovery of a RuBisCO-like Protein that Functions as an Oxygenase in the Novel d-Hamamelose Pathway. Biotechnol. Bioprocess Eng. 23(5):490–99
Kono T, Mehrotra S, Endo C, Kizu N, Matusda M, et al. (2017). A RuBisCO-mediated carbon metabolic pathway in methanogenic archaea. Nat. Commun. 8:14007
Kreel NE, Tabita FR. (2007). Substitutions at methionine 295 of Archaeoglobus fulgidus ribulose-1,5-bisphosphate carboxylase/oxygenase affect oxygen binding and CO2/O2 specificity. J. Biol. Chem. 282(2):1341–51
Liu D, Ramya RCS, Mueller-Cajar O. (2017). Surveying the expanding prokaryotic Rubisco multiverse. FEMS Microbiol. Lett. 364(16):
Madigan, M. T. et al. (2020): Brock Mikrobiologie. 15. Auflage, Pearson-Verlag
Martins, BM. et al. (2004): Crystal structure of 4-hydroxybutyryl-CoA dehydratase: Radical catalysis involving a [4Fe–4S] cluster and flavin. PNAS 101(44); 15645–15649
Miyazaki JH, Yang SF (1987): The methionine salvage pathway in relation to ethylene and polyamine biosynthesis. Phys. Plant. 69(2): 366–370
Morell MK, Paul K, Kane HJ, Andrews TJ (1992). Rubisco: Maladapted or Misunderstood. Aust. J. Bot. 40(5):431–41
Munk, K. (2008): Taschenlehrbuch Biologie: Botanik. Stuttgart: Georg Thieme Verlag.
Nelson, D.; Cox, M. (2010): Lehninger Biochemie. Wiesbaden: Springer Berlin Heidelberg.
Nisbet EG, Grassineau NV, Howe CJ, Abell PI, Regelous M, Nisbet RER. (2007). The age of Rubisco: the evolution of oxygenic photosynthesis. Geobiology 5: 311–335.
Osborne CP, Beerling DJ (2006). Nature’s green revolution: the remarkable evolutionary rise of C4 plants. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 361 (1465): 173–94.
Osborne CP, Freckleton RP (2009). Ecological selection pressures for C4 photosynthesis in the grasses. Proceedings. Biological Sciences. 276 (1663): 1753–60.
Paoli GC, Soyer F, Shively J, Tabita FR. (1998). Rhodobacter capsulatus genes encoding form I ribulose-1,5-bisphosphate carboxylase/oxygenase (cbbLS) and neighbouring genes were acquired by a horizontal gene transfer. Microbiology 144: 219–227. pmid:9467914
Pirkov I, Norbeck J, Gustafsson L, Albers E (2008): A complete inventory of all enzymes in the eukaryotic methionine salvage pathway. FEBS J. 275 (16): 4111–20.
Prywes N, Phillips NR, Tuck OT, Valentin-Alvarado LE, Savage DF (2023). Rubisco Function, Evolution, and Engineering. Annu Rev Biochem. 92:385-410.
Raven JA (2009): Contributions of anoxygenic and oxygenic phototrophy and chemolithotrophy to carbon and oxygen fluxes in aquatic environments. Aquat. Microb. Ecol. 56:177–92
Sadava, D., et al. (2019): Purves Biologie. Berlin Heidelberg New York: Springer-Verlag.
Sage RF (2004). The evolution of C4 photosynthesis. New Phytologist. 161 (2): 341–370
Sage RF (2017). A portrait of the C4 photosynthetic family on the 50th anniversary of its discovery: species number, evolutionary lineages, and Hall of Fame. Journal of Experimental Botany 68(2): e11–e28.
Sage R, Monson R (1999). „7“. C4 Plant Biology. pp. 228–229. ISBN 978-0-12-614440-6.
Sage R, Monson R, Ehleringer J, Adachi S, Pearcy R (2018). Some like it hot: the physiological ecology of C4 plant evolution. Oecologia. 187.
Sato T, Atomi H, Imanaka T. (2007). Archaeal type III RuBisCOs function in a pathway for AMP metabolism. Science. 315(5814):1003–6
Schwender J, Goffman F, Ohlrogge JB, Shachar-Hill Y. (2004). Rubisco without the Calvin cycle improves the carbon efficiency of developing green seeds. Nature. 432(7018):779–82
Shih P, Ward LM, Fischer W (2017). Evolution of the 3-hydroxypropionate bicycle and recent transfer of anoxygenic photosynthesis into the Chloroflexi. Proceedings of the National Academy of Sciences. 114. 201710798.
Sinclair TR, Rufty TW, Lewis RS. (2019). Increasing Photosynthesis: Unlikely Solution For World Food Problem. Trends Plant Sci. 0(0)
Strauss G, Fuchs G (1993). Enzymes of a novel autotrophic CO2 fixation pathway in the phototrophic bacterium Chloroflexus aurantiacus, the 3-hydroxypropionate cycle. European Journal of Biochemistry. 215 (3): 633–43.
Stryer L, Berg JM, Tymoczko JL (2002). Chapter 20: The Calvin Cycle and the Pentose Phosphate Pathway“. Biochemistry (5th ed.). San Francisco: W.H. Freeman.
Tabita FR, Hanson TE, Li H, Satagopan S, Singh J, Chan S. (2007). Function, structure, and evolution of the RubisCO-like proteins and their RubisCO homologs. Microbiol. Mol. Biol. Rev. 71(4):576–99
Tabita FR, Satagopan S, Hanson TE, Kreel NE, Scott SS. (2008). Distinct form I, II, III, and IV Rubisco proteins from the three kingdoms of life provide clues about Rubisco evolution and structure/function relationships. J. Exp. Bot. 59(7):1515–24
Thauer RK (2007): Microbiology. A fifth pathway of carbon fixation. Science318 (5857), 1782–1783.
Vitlin Gruber A, Feiz L (2018). „Rubisco Assembly in the Chloroplast“. Frontiers in Molecular Biosciences. 5: 24. doi:10.3389/fmolb.2018.0002
Yoon M, Putterill JJ, Ross GS, Laing WA (2001). Determination of the relative expression levels of rubisco small subunit genes in Arabidopsis by rapid amplification of cDNA ends. Analytical Biochemistry. 291 (2): 237–244.
Warke MR, Di Rocco T, Zerkle AL, Lepland A, Prave AR, Martin AP, et al. (2020). The Great Oxidation Event preceded a Paleoproterozoic “snowball Earth”. Proc Natl Acad Sci U S A. 117: 13314–13320.
Watson GM, Yu JP, Tabita FR. (1999). Unusual ribulose 1,5-bisphosphate carboxylase/oxygenase of anoxic Archaea. J. Bacteriol. 181(5):1569–75
Weiss, M. C., Sousa, F. L., Mrnjavac, N., Neukirchen, S., Roettger, M., Nelson-Sathi, S., et al. (2016): The physiology and habitat of the last universal common ancestor. Nat. Microbiol. 1:16116.
Whitney SM, Shaw DC, Yellowlees D. (1995). Evidence that some dinoflagellates contain a ribulose-1,5-bisphosphate carboxylase/oxygenase related to that of the alpha-proteobacteria. Proc. Biol. Sci. 259(1356):271–75
Williams BP, Johnston IG, Covshoff S, Hibberd JM (2013). Phenotypic landscape inference reveals multiple evolutionary paths to C4 photosynthesis. eLife. eLife Sciences Publications. 2: e00961.
Wray JW, Abeles RH (1995): The methionine salvage pathway in Klebsiella pneumoniae and rat liver. Identification and characterization of two novel dioxygenases. In: J. Biol. Chem. 270(7): 3147–53
Wrighton KC, Castelle CJ, Varaljay VA, Satagopan S, Brown CT, et al. (2016). RubisCO of a nucleoside pathway known from Archaea is found in diverse uncultivated phyla in bacteria. ISME J. 10(11):2702–14
Zarzycki J, Brecht V, Müller M, Fuchs G (2009). Identifying the missing steps of the autotrophic 3-hydroxypropionate CO2 fixation cycle in Chloroflexus aurantiacus. Proceedings of the National Academy of Sciences of the United States of America. 106 (50): 21317–22.
