
In the last episode, we learned how to recognize a dinosaur by its apomorphies. If you don’t know these yet, take another look at the corresponding episode. Dinosaurs are part of the Dinosauromorpha clade, which in turn is part of the Ornithodira clade. The sister group of the Dinosauromorpha are the Pterosauromorpha, to which the pterosaurs belong. Ornithodira are part of the Avemetatarsalia and these, together with the Pseudosuchia or Crurotarsi, the crocodilian relatives, are part of the Archosauria.
pdf file
Origin of Dinosaurs
When did the first dinosaurs appear? The fossils of the oldest undoubted dinosaurs, Eoraptor and Herrerasaurus, come from the Ischigualasto Formation in Argentina, which is around 230 million years old, i.e. from the late Triassic (Fig. 1). Eoraptor is considered the most primitive representative and probably looked very similar to the common ancestor of all dinosaurs (Sereno 1999, Sereno et al. 1988, 1993, 2013, Sereno & Novas 1992, Sterling et al. 2012, Reig 1963, Novas et al. 2021). This means that the first dinosaurs were probably small, bipedal carnivores.
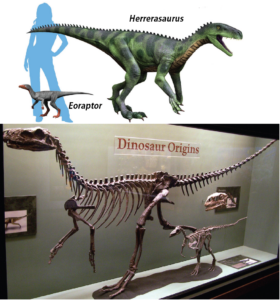
Fig. 1: Eoraptor and Herrerasaurus are one of the earlies dinosaurs
In East Africa, the fossil Nyasasaurus was discovered, which lived 245 million years ago and could be either the earliest dinosaur or the closest known relative of the dinosaurs (Nesbitt et al. 2013, 2020, Novas et al. 2021). If the latter is the case, it belongs to the clade Dinosauriformes within Dinosauromorpha, which would be the sister group of dinosaurs. Since only few and fragmented fossils of Nyasasaurus have been found, only a few morphological features of the animals can be determined, which makes its exact classification in the phylogenetic tree uncertain. The Dinosauromorpha do have some of the characteristics of the „real dinosaurs“, but not all of them. To the non-specialist, they look superficially like dinosaurs, but they are not quite dinosaurs. Some smaller genera such as Lagosuchus, Marasuchus and Silesaurus belong to this group (Angolin & Martin 2019, Dzik 2003, Sereno et al. 1994, Fig. 2). Dinosaurs with their own apomorphies then evolved from these or similar forms. If some Dinosauromorpha are more similar to dinosaurs than others, but still not dinosaurs, they are simply (together with the dinosaurs) Dinosauriformes.
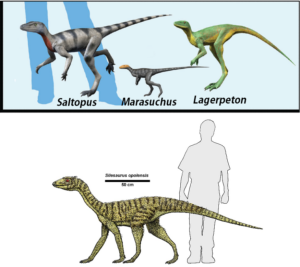
Fig. 2: Dinosauromorpha
For a better understanding, let’s take a closer look at one representative: Silesaurus. It lived 230 million years ago, was about 2 meters long, had four long slender legs and a graceful physique. Although outwardly very similar to a dinosaur, it still had some significant differences to the „real dinosaurs“. Although its thigh bones ran vertically, they were connected to the side of the hip, unlike those of dinosaurs. And because the joint head of the femur in Silesaurus was neither inclined nor fitted into an open joint socket – typical of dinosaurs – and the legs were not under the body, it was not a true dinosaur. Although Silesaurus walked on four legs, the front legs were used for support rather than locomotion. It was only a small step before the first dinosaurs balanced their bodies so that they could move on two legs. This offered the advantage of being able to run faster. At the same time, it opened up new possibilities for the hands, for example, they could reach for food. This was the dinosaurs‘ recipe for success and the reason why they were able to prevail over the other dinosauromorphs at the end of the Triassic (Schmitt 2023).
Saurischia – Ornithischia – Ornithoscelida
Starting with early dinosaurs such as Eoraptor, what was the evolutionary history of dinosaurs? In the last article, we learned that there are two main clades within the dinosaurs: the lizard-hipped dinosaurs (Saurischia), which unite the carnivorous theropods and the long-necked sauropodomorphs, and the bird-hipped dinosaurs (Ornithischia), which include all other herbivorous dinosaurs such as Triceratops, Stegosaurus and Iguanodon (Fig. 3). Despite the term „bird-hipped dinosaurs“, birds do not belong to this group, but to the lizard-hipped dinosaurs, as they evolved from theropods (Fastovsky & Weishampel 2021, Holtz 2007, Paul 2016, Prothero 2017, 2021, 2022, Prothero & Dott 2004, Brusatte 2019, Schweitzer et al. 2021).
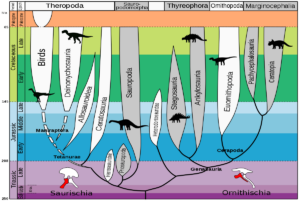
Fig. 3: Dinosaur phylogenetic tree
The similarity between the pelvis of bird-hipped dinosaurs and birds lies in the position of the pubis. Unlike in lizard-hipped dinosaurs, the pubis in bird-hipped dinosaurs does not point downwards and slightly forwards towards the head, but consists of a narrow, rod-shaped bone that lies lengthways next to the ischium and points backwards. In birds, the pubis is completely displaced backwards (Fig. 4).
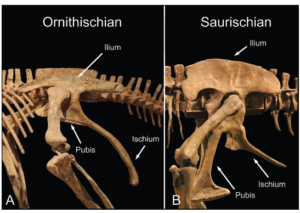
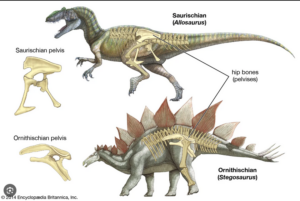
Fig. 4: Ornithischia and Saurischia
In a detailed phylogenetic study from 2017, based on 74 taxa and 457 features, the palaeontologists Matthew G. Baron, David B. Norman and Paul M. Barrett propose a radically new systematics of dinosaurs (Baron et al. 2017a, Padian 2017). Accordingly, Ornithischia and Theropoda are united as sister groups in the clade Ornithoscelida, while the Sauropodomorpha together with the Herrerasauridae form the newly defined clade Saurischia (Fig. 5). The Ornithoscelida hypothesis was challenged by an international team of researchers in November 2017 after the original anatomical dataset of Baron et al. (2017a) was revised. This revision resulted in the traditional model in which Ornithischia and Saurischia were recovered as sister taxa. However, this traditional tree was only weakly supported and was not statistically significantly different from the alternative Ornithoscelida hypothesis (Langer et al. 2017). With only minor adjustments subsequently made by Baron and colleagues, Ornithoscelida was again favored over the traditional model (Baron et al. 2017b).
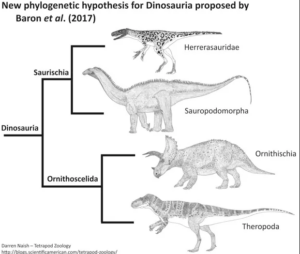
Fig. 5: Ornithoscelida
Since the debate about the validity of the Ornithoscelida concept is still ongoing and most dinosaur textbooks follow the traditional concept, we will stick to the traditional concept. However, the relationships within the Ornithischia, Theropoda and Sauropodomorpha are compatible in both concepts.
Considering how rare dinosaur fossils are, especially compared to marine invertebrates, it is remarkable that we not only have quite a large diversity of dinosaur genera – about 1000 genera described – but also have enough specimens to show that we have transitions between almost all major dinosaur groups, as well as many other remarkable fossils showing other types of transitions, such as carnivorous dinosaurs that became herbivores. In this post, we will first look at those dinosaurs that have nothing to do with birds.
Evolutionary transitions in Sauropodomorpha
The gigantic and long-necked sauropods are among the most breathtaking dinosaurs. Among them were not only giants such as Diplodocus, Brachiosaurus or Argentinosaurus, but also dwarf forms such as Europasaurus (Fig. 6). The reason for the gigantism of sauropods has already been discussed in a separate article by me (in German) (Sander & Clauss 2008, Sander et al. 2011, 2013, Klein et al. 2013, Clauss et al. 2013, Christian et al. 2013, Stevens 2013, Taylor & Wedel 2013a,b, Sellers et al. 2013, Wedel & Taylor 2013, Fastovsky & Weishampel 2021, Holtz 2007, Paul 2016, Prothero 2022, Schweitzer et al. 2021, Manz 2022). Are there actually evolutionary transitions here? Yes!

Fig. 6: Size comparison of the sauropods Argentinosaurus and Europasaurus
Almost every book on dinosaurs (e.g. Fastovsky & Weishampel 2021, Holtz 2007, Paul 2016, Schweitzer et al. 2021) depicts a group of creatures from the Triassic called prosauropods. Their very name (translated as „before the sauropods“) indicates that they are original relatives of the larger sauropods. The best known of these is Plateosaurus from the Triassic in Germany, but there are a dozen other genera that have been found in Triassic strata around the world (Fig. 7 – 9). Most of these genera were only about 5-8 meters long, only a quarter the size of the giant Jurassic sauropods, but larger than their ancestors. They have the beginnings of a long neck and a long tail, but not yet the incredible dimensions of the giant sauropods. The limbs are classically sauropod in the construction of the fingers and toes, but not as robust, and the forelimbs are long and delicate enough that they could switch between quadrupedal walking and rearing up on their hind legs in a bipedal stance to use their hands. It was only when sauropods reached enormous size that they were forced to walk exclusively on all fours, and their limbs also became much more massive to support their enormous body weight.

Fig. 7: Plateosaurus and Massospondylus
Fig. 8: Prosauropods: A) Guaibasaurus, (B) Panphagia, (C) Saturnalia, (D) Pantydraco, (E) Bagualosaurus, (F) Thecodontosaurus
Fig. 9: The prosauropod Plateosaurus compared to the sauropods (Brachiosaurus and Barosaurus). Like other basal sauropodomorphs, Plateosaurus had an elongated neck and was bipedal. All sauropodomorphs have at least 10 cervical vertebrae (Plateosaurus does not show cervical vertebra No. 1 in the figure above left). The number of cervical vertebrae has increased in sauropods in the course of evolution; Barosaurus had 16. In the course of the evolution of sauropodomorphs, gigantism led to a reorganization of the forelimbs (illustrated by the example of Brachiosaurus, on the right in the picture). The humerus, radius and ulna as well as the metacarpal bones are arranged in a vertical column. In addition, the carpal bones and fingers have been reduced. A sauropod dorsal vertebra with labeled lamina and fossa can also be seen. These structures are the result of postcranial pneumatization, a process in which extensions of the lung penetrate the bone.


Anchisaurus from the Triassic of Connecticut, Arizona and South Africa was even more primitive than Plateosaurus (Yates 2004, Prothero 2017, Fig. 10). It was only 2.5 meters long, only slightly larger than a human, and had an even shorter neck and tail and even more delicate limbs and feet. In fact, it is the perfect transitional form between the lizard-like early dinosaurs such as the much smaller Lagosuchus and larger forms such as Plateosaurus. Despite its outward appearance, its skull bears all the characteristic features of sauropods, and it already shows many of the specializations in the vertebrae and especially in the hands and feet that will later characterize sauropods. The early dinosaurs were primarily bipedal, but Anchisaurus appears to have been capable of both postures, and Plateosaurus was even heavier and more quadrupedal.

Fig. 10: Anchisaurus
From the earliest known dinosaur Eoraptor to Anchisaurus and Plateosaurus to the larger sauropods, there is not only a smooth increase in size, but also a smooth transition in anatomical features and body posture from biped to quadruped (Fig. 11).
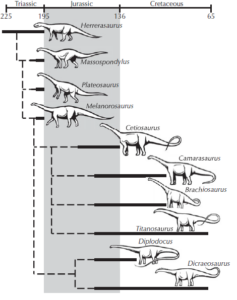
Fig. 11: phylogenetic tree of sauropods. The prosauropods were transitional forms between smaller primitive bipedal dinosaurs such as Eoraptor and Herrerasaurus and the sauropods.
Evolutionary transitions in non-avian theropods
The other main branch of lizard-hipped dinosaurs were the theropods, predatory dinosaurs, which include the famous Tyrannosaurus rex, but there are dozens of different genera and species in all shapes and sizes. However, the early theropods were quite inconspicuous and are actually excellent transitional forms. They ranged in length from just 70 centimeters (like Compsognathus) to 3 meters (like Coelophysis), meaning most were about the size of a chicken to the size of an adult human. Unlike their larger theropod descendants, they were lightly built, with small heads, long necks and slender, graceful limbs and tails. Nevertheless, their skulls and especially their hands (with their unique combination of only three fingers: thumb, index and middle finger) and feet had all the anatomical specializations found in later theropods.
From original theropods like Coelophysis (Fig. 12), we can trace the theropod lineage even further back to Eoraptor, Staurikosaurus and Herrerasaurus, which are built similarly to Coelophysis, but do not yet show all the distinct specializations of theropods (Fastovsky & Weishampel 2021, Holtz 2007, Paul 2016, Prothero 2017, 2021, 2022, Prothero & Dott 2004, Brusette 2012, 2019, Schweitzer et al. 2021).
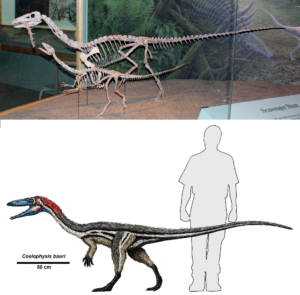
Fig. 12: Coelophysis
To a casual observer they look very similar, but to a paleontologist with anatomical training the differences are clear. Eoraptor, Staurikosaurus and Herrerasaurus lack the highly specialized three-fingered hand (some still had the full five fingers, Fig. 13), the relatively unspecialized vertebrae (both sauropod and theropod vertebrae are highly specialized and distinct), a sliding temporomandibular joint, the fully curved carnivore teeth, and the ankle and foot modifications typical of theropods found in Coelophysis and more advanced theropods (Schwartz et al. 1994, Rinehart et al. 2004, 2007, 2009, Gay 2001, Downs 2000, Barta et al. 2018, Prothero 2017, Ezcurra 2007) Finally, we can trace genera such as Eoraptor, Staurikosaurus and Herrerasaurus back to even more primitive, non-dinosaur-like archosaurs such as Euparkeria, which superficially resemble the earliest dinosaurs but lack the unique specializations that all dinosaurs have, such as the open hip joint. such as the open acetabulum or the characteristic features of the skull.
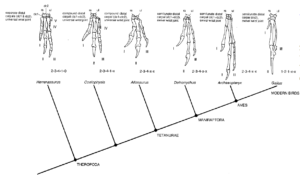
Fig. 13: The left hand of selected theropods in a phylogenetic context shows the pattern of finger reduction and the development of the wrist from a universal joint to a swivel joint. The problem of counting fingers in theropods (I-II-III) and birds (II-III-IV) will be discussed separately in another article.
So we can trace both the sauropods (via the prosauropods) and the theropods (via the primitive genera such as Coelophysis) back to a common Saurischian ancestor in the likes of Eoraptor, Staurikosaurus and Herrerasaurus, and from there to more primitive archosaurs that were not dinosaurs, such as Euparkeria. You couldn’t ask for a more beautiful series of transitional forms. Obviously, creationists have never heard of any of these forms. And there’s a cool final twist to this story of predatory dinosaurs. In 2005, Jim Kirkland and others announced the discovery of a remarkable new fossil called Falcarius utahensis from the Jurassic of Utah (Kirkland et al. 2005, Zanno 2010a,b, Zanno & Erickson 2006, Fig. 14). This strange creature belongs to an even stranger group, the therizinosaurs, whose exact position within the dinosaurs has long been disputed. These animals have many of the features of theropods like Velociraptor, including long clawed fingers on their hands and a long neck and tail for balancing. But they had toothless beaks and were apparently herbivores. In recent years, there has been a consensus that therizinosaurs were indeed theropods that had somehow reverted to herbivory. The discovery of Falcarius provided the „missing link“ in this dietary change, as it retains many features of „raptor“ dinosaurs, but is the most primitive therizinosaur with a toothless herbivorous beak. This made it a classic transitional form, not only linking therizinosaurs to raptors anatomically, but also showing how they made the remarkable transition from carnivore back to raptor.

Fig. 14: Falcarius utahensis
Evolutionary transitions in Ornithischia
The other main branch of dinosaurs is the Ornithischia, which includes almost all herbivorous dinosaurs (except the sauropods): the duckbills, the iguanodonts, the armored ankylosaurs, the spiny stegosaurs, the thick-headed pachycephalosaurs and the horned ceratopsians. The earliest ornithischians include primitive Triassic forms such as Lesothosaurus, Fabrosaurus and Heterodontosaurus, small bipedal dinosaurs that superficially look like Eoraptor or Coelophysis (Fig. 15-17). On closer inspection, however, they have all the characteristics of Ornithischia: part or all of the pubis at the hip is rotated backwards parallel to the ischium; the molars are deeply set into the jaw, suggesting that they had cheeks to hold their food in their mouths while chewing; and they have a unique extra bone at the tip of the lower jaw known as the praedentary (Fig. 18). All of these features are unique to the Ornithischia, but we can see that the transitional forms such as Heterodontosaurus already had them in the Triassic, while they still resembled the other primitive dinosaurs of the time (Fastovsky & Weishampel 2021, Holtz 2007, Paul 2016, Prothero 2017, 2022, Schweitzer et al. 2021).
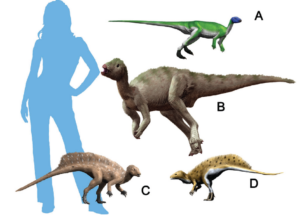
Fig. 15: early Ornithischia. (A) Lesothosaurus, (B) Hypsilophodon, (C) Heterodontosaurus, (D) Tianyulong.
Fig. 16:early Ornithischia. (A) Hypsilophodon, (B) Leaellynasaura, (C) Orodromeus, (D) Kulindadromeus.
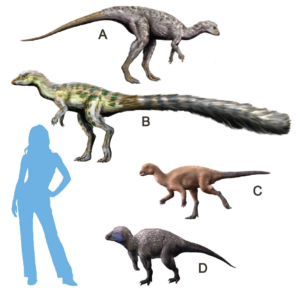

Fig. 17: Skelett des Heterodontosaurus

Fig. 18: Synapomorphies of the Ornithischia. Left: Pubic bone of a Lambeosaurus (cast). In this and other Ornithischia, the pubis points backwards (arrows show the caudal end of the dinosaur, almost parallel to the ischium. Right: Skull (cast) of a Maiasaura, with ornithischian features marked, including the praedentary and a mandibular joint that lies below the row of teeth of the upper jaw.
Among the most popular ornithischians is Triceratops, one of the last horned dinosaurs or ceratopsians. Ceratopsians are another classic case of transitional forms between highly specialized forms, such as the horned and frilled ceratopsians, and much more primitive forms that resemble the common ancestor with other dinosaur clades (Figs. 19 and 20).

Fig. 19: Ceratopsia. (E) Diabloceratops, (F) Pachyrhinosaurus, (G) Styracosaurus, (H) Agujaceratops, (I) Chasmosaurus, (J) Pentaceratops, (K) Triceratops

Fig. 20: early Ceratopsia. (A) Yinlong, (B) Psittacosaurus, (C) Protoceratops, (D) Leptoceratops
All genera with horns can be traced back to the very well-known Protoceratops, which has the frill and the characteristic bones in the beak and lower jaw, but lacks horns (Fastovsky & Weishampel 2021, Holtz 2007, Paul 2016, Weishampel et al. 1990, Granger & Gregory 1923, Fig. 20 & 21). Creationists such as the US guru Duane Gish, however, do not seem to understand the position of Protoceratops as a transitionial form between different Ceratopsians (Gish 1995). He cites an out-of-context quote from the first edition of Weishampel et al (1990) about the distinctiveness of Protoceratopsidae (completely overlooking the fact that this doesn’t make them any less of a good transitional form), and he also mentions that they occur in the Late Cretaceous and therefore cannot be ancestral. I already mentioned in my post on the topic of transitional forms (in German) that paleontologists are not looking for ancestors, but for sister groups that demonstrate anatomical transitions. Due to the rarity of fossilization, it is not to be expected that the direct ancestor of an evolutionary line will be discovered. Secondly, Protoceratops occurred in the early Late Cretaceous, millions of years before all its putative descendants among the horned Ceratopsia in the late Late Cretaceous. Moreover, we have even better transitional forms.

Fig. 21: Protoceratops
Bagaceratops has a slightly smaller frill and beak compared to Protoceratops, and its body is not fully quadrupedal (Maryanska & Osmolska 1975, Czepinsky 2019, Fig. 22). Archaeoceratops has an even smaller frill and beak and a much lighter body that is most likely bipedal (Tang et al. 2001, Jin et al. 2009, Fig. 23). Psittacosaurus, so named because it had a parrot-like beak, has the beginnings of a frill over the neck and is much lighter in build with a bipedal, graceful skeleton instead of the heavier skeleton of Protoceratops. Psittacosaurus shows not only the transition from a frillless skull to one with a small frill to the larger frill of Protoceratops, but also the transition from a light bipedal body (typical of almost all basal dinosaurs) to the heavier quadrupedal body of the more specialized groups (Osborn 1923, Coombs 1982, Sereno 2010, Mayr et al. 2002, Dalton 2001, Lingham-Soliar 2001, Vinther et al. 2016, 2021, Figs. 20 & 24).
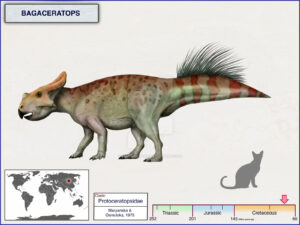
Fig. 22: Bagaceratops
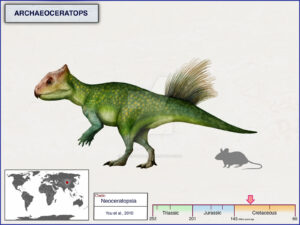
Fig. 23: Archaeoceratops

Fig. 24: Psittacosaurus.
The most amazing transitional fossil in this sequence was finally discovered in 2004 with Yinlong from the much earlier Late Jurassic strata in China (Xu et al. 2006, Figs. 20 & 25). The name Yinlong means „hidden dragon“ in Mandarin, in reference to the popular movie „Crouching Tiger, Hidden Dragon“, part of which was filmed near the site. Yinlong consists of a well-preserved skeleton of a bipedal dinosaur, not too different in proportions from Psittacosaurus. At the tip of its upper beak it has the rostral bone typical of ceratopsians. However, its skullcap has a unique arrangement of bones found in the thick-skulled Pachycephalosaurs, which are known to have a thick dome of bone in their skulls. Paleontologists have long argued that Ceratopsia and Pachycephalosaurs are closely related because they both have a bony crest at the back of the skull. Both are grouped together as „Marginocephalia“ (Fastovsky & Weishampel 2021, Holtz 2007, Paul 2016, Prothero 2017, 2022, Schweitzer et al. 2021) (Fig. 26). But with Yinlong we have a beautiful transitional fossil that shows characteristics of both ceratopsians and pachycephalosaurs before both families split.
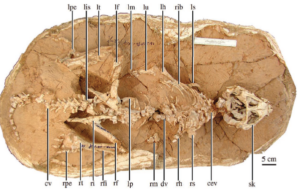
Fig. 25: Fossil of Yinlong
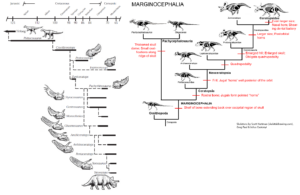
Fig. 26. Evolution of Marginocephalia
The transitions of the armored anklyosaurs and stegosaurs are also worth mentioning. Both families are grouped together as Thyreophora and are characterized by apomorphies unique to this group, such as armor plates lined up along the body, relatively small brains and a quadrupedal posture with hind limbs that are longer than the forelimbs (Fastovsky & Weishampel 2021, Holtz 2007, Paul 2016, Prothero 2017, 2022, Schweitzer et al. 2021, Fig. 27).
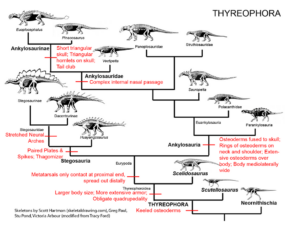
Fig. 27: Thyreophora
For example, the basal anklyosaurs – the nodosaurs – still have very limited armor, as do the basal stegosaurs, such as Scelidosaurus (Fig. 28). We can trace all these forms back to genera such as Scutellosaurus, a small, approximately 1.2 m long dinosaur from the Lower Jurassic 200 million years ago (Fig. 29). It had even longer hind legs and could move on both two and four legs. Its body was covered with small horny scales, which were probably attached in rows to the back and flanks of the animal (Norman 2020, 2021, Norman et al. 2004, Gierlinski 1999, Maidment & Porro 2010, Martill et al. 2000, Carpenter 2001, Thompson et al. 2011, Colbert 1981, Prothero 2017, 2021, Fastovsky & Wieshampel 2021, Schweitzer et al. 2021).
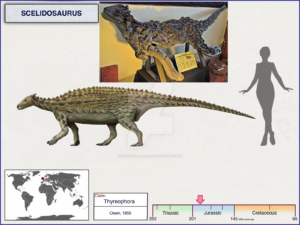
Fig. 28: Scelidosaurus
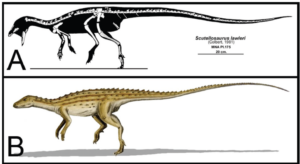
Fig. 29: Scutellosaurus
We can trace all Ornithischia back to the earliest forms, such as Heterodontosaurus, Fabrosaurus or Lesothosaurus. These genera in turn looked very similar in their external features to Eoraptor and Herrerasaurus, the earliest dinosaurs, except for a few subtle differences, such as the presence of a praedentary at the tip of the lower jaw and a primitive ornithischian hip structure. If one reads the literature carefully and without bias, these transitions are obvious.
We could go on endlessly debunking creationist falsehoods about dinosaurs. Anyone with even a moderate interest in the subject need only buy a dinosaur book – sometimes a children’s book will do! – and look at the beautiful transitional forms for almost every group that creationists deny. But this should be enough for now. If we can find transitions between such different dinosaurs, the transition between birds and theropod dinosaurs is even more exciting. And we will get to know this in the next episode.
Literature
Agnolin, F. L., Ezcurra, M. D. (2019): The Validity of Lagosuchus Talampayensis Romer, 1971 (Archosauria, Dinosauriformes), from the Late Triassic of Argentina. Breviora, 565(1):1-21
Baron, M. G., Norman, D. B., Barrett, P. M. (2017a): A new hypothesis of dinosaur relationships and early dinosaur evolution. Nature 543:501–506.
Baron, M. G., Norman, D. B., Barrett, P. M. (2017b). Baron et al. reply. Nature 551(7678): E4–E5.
Barta, D. E., Nesbitt, S. J., Norell, M. A. (2018): The evolution of the manus of early theropod dinosaurs is characterized by high inter‐ and intraspecific variation. Journal of Anatomy. 232 (1): 80–104.
Brusatte, S. L. (2019): The Rise and Fall of the Dinosaurs: A New History of a Lost World. William Morrow, NY
Carpenter, K. (2001): Phylogenetic Analysis of Ankylosauria. In Carpenter, Kenneth (ed.). The Armored Dinosaurs. Indiana University Press. pp. 455–480.
Clauss, M., Steuer, P., Müller, D. W. H., Codron, D., Hummel, J. (2013): Herbivory and Body Size: Allometries of Diet Quality and Gastrointestinal Physiology, and Implications for Herbivore Ecology and Dinosaur Gigantism. PLoS ONE 8(10): e68714.
Christian, A., Peng, G., Sekiya, T., Ye, Y., Wulf, M. G., Steuer, T. (2013): Biomechanical Reconstructions and Selective Advantages of Neck Poses and Feeding Strategies of Sauropods with the Example of Mamenchisaurus youngi. PLoS ONE 8(10): e71172.
Colbert, E. H. (1981): A primitive ornithischian dinosaur from the Kayenta Formation of Arizona. Museum of Northern Arizona Bulletin 53:1-61
Coombs, W. P. (1982): Juvenile specimens of the ornithischian dinosaur Psittacosaurus. Palaeontology. 25: 89–107
Czepiński, Ł. (2019): Ontogeny and variation of a protoceratopsid dinosaur Bagaceratops rozhdestvenskyi from the Late Cretaceous of the Gobi Desert. Historical Biology. 32 (10): 1394–1421.
Dalton, R. (2001): Wandering Chinese fossil turns up at museum. Nature 414: 571.
Downs, A. (2000): Coelophysis bauri and Syntarsus rhodesiensis compared, with comments on the preparation and preservation of fossils from the Ghost Ranch Coelophysis quarry: In: Lucas, S.G., Heckert, A.B. (eds.). „Dinosaurs of New Mexico“. New Mexico Museum of Natural History Bulletin. 17: 33–37
Dzik, J. (2003): A beaked herbivorous archosaur with dinosaur affinities from the early Late Triassic of Poland. Journal of Vertebrate Paleontology. 23 (3): 556–574.
Ezcurra, M. D. (2007): The cranial anatomy of the coelophysoid theropod Zupaysaurus rougieri from the Upper Triassic of Argentina. Historical Biology. 19 (2): 185–202.
Fastovsky, D. E., Weishampel, D. B. (2021): Dinosaurs: A Concise Natural History (4th ed.). Cambridge university Press, Cambridge.
Gay, R.J. (2001): An unusual adaptation in the caudal vertebrae of Coelophysis bauri (Dinosauria: Theropoda). PaleoBios. 21: 55.
Gierliński, G. (1999): Tracks of a large thyreophoran from the Early Jurassic of Poland. Acta Palaeontologica Polonica. 44.
Gish, D. (1995): Evolution, the Fossils Still Say NO! San Diego, Calif.: Creation-Life.
Granger, W. W., Gregory, W. K. (1923): Protoceratops andrewsi, a pre-ceratopsian dinosaur from Mongolia. American Museum of Natural History Novitates (72): 1−9.
Holtz, T. R. (2007): Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages
Jin L., Chen J., Zan S., Godefroit, P. (2009): A New Basal Neoceratopsian Dinosaur from the Middle Cretaceous of Jilin Province, China. Acta Geologica Sinica. 83 (2): 200–206.
Kirkland, J. I., Zanno, L. E., Sampson, S. D., Clark, J. M., DeBlieux, D. D. (2005): A primitive therizinosauroid dinosaur from the Early Cretaceous of Utah. Nature. 435 (7038): 84–87.
Klein, N., Remes, K., Gee, C. T., Sander, P. M. (2011): Biology of the Sauropod Dinosaurs: Understanding the Life of Giants. Bloomington: Indiana University Press
Langer, M. C., Ezcurra, M. D., Rauhut, O. W. M., Benton, M. J., Knoll, F., McPhee, B. W., Novas, F. E., Pol, D., Brusatte, S. L. (2017): Untangling the dinosaur family tree. Nature551(7678): E1–E3.
Lingham-Soliar, T. (2008): A unique cross section through the skin of the dinosaur Psittacosaurus from China showing a complex fibre architecture. Proceedings of the Royal Society B: Biological Sciences. 275 (1636): 775–780.
Maidment, S. C. R., Porro, L.B. (2010): Homology of the palpebral and origin of the supraorbital ossifications in ornithischian dinosaurs. Lethaia, 43: 95-111
Manz, A. (2022): Sauropoden – dem Gigantismus auf der Spur https://www.scinexx.de/dossier/sauropoden-dem-gigantismus-auf-der-spur/
Martill, D. M., Batten, D. J., Loydell, D. K. (2000): A New Specimen of the Thyreophoran Dinosaur cf. Scelidosaurus with Soft Tissue Preservation. Palaeontology 43(3): 549-559.
Maryańska, T., Osmólska, H. (1975): Protoceratopsidae (Dinosauria) of Asia. Palaeontologia Polonica. 33: 134−143.
Mayr, G., Peters, S. D., Plodowski, G., Vogel, O. (2002): Bristle-like integumentary structures at the tail of the horned dinosaur Psittacosaurus. Naturwissenschaften. 89 (8): 361–365.
Nesbitt, S. J., Barrett, P. M., Werning, S., Sidor, C. A., Charig, A. J. (2013): The oldest dinosaur? A Middle Triassic dinosauriform from Tanzania. Biol. Lett. 9 (1): 20120949.
Nesbitt, S, Butler, R, Ezcurra, M, Charig, A, Barrett, P (2020): The anatomy of Teleocrater Rhadinus, an early avemetatarsalian from the lower portion of the Lifua Member of the Manda Beds (Middle Triassic) (project). MorphoBank datasets.
Norman, D. B. (2020): Scelidosaurus harrisonii from the Early Jurassic of Dorset, England: cranial anatomy. Zoological Journal of the Linnean Society. 188 (1): 1–81.
Norman, D. B. (2021): Scelidosaurus harrisonii (Dinosauria: Ornithischia) from the Early Jurassic of Dorset, England: biology and phylogenetic relationships. Zoological Journal of the Linnean Society. 191(1):1–86.
Norman, D. B., Witmer, L. M., Weishampel, D. B. (2004): Basal Thyreophora. In Weishampel, D.B., Dodson, P., and Osmólska, H. (ed.). The Dinosauria, 2nd Edition. University of Californian Press. pp. 335–342
Novas, F. E., Agnolin, F. L., Ezcurra, M. D., Temp Müller, R., Martinelli, A. G., Langer, M. C. (2021): Review of the fossil record of early dinosaurs from South America, and its phylogenetic implications. Journal of South American Earth Sciences. 110: 103341.
Osborn, H. F. (1923): Two Lower Cretaceous dinosaurs of Mongolia. American Museum Novitates (13): 1–10.
Padian, K. (2017): Palaeontology: Dividing the dinosaurs. Nature 543:494–495.
Paul, G. S. (2016): The Princeton Fieldguide to Dinosaurs. 2nd Edition. Princeton University Press
Prothero, D. (2017): Evolution – What the Fossils say and why it matters. Second edition. New York: Columbia University Press
Prothero, D. (2021): The Evolving Earth. Oxford University Press
Prothero, D. (2022): Vertebrate Evolution. From Origins to Dinosaurs and beyond. CRC Press, Kapitel 11, 12, 14
Prothero, D. & Dott, (2004): Evolution of the Earth, Seventh edition. McGrawHill
Reig, O.A. (1963): La presencia de dinosaurios saurisquios en los „Estratos de Ischigualasto“ (Mesotriásico Superior) de las provincias de San Juan y La Rioja (República Argentina). Ameghiniana 3 (1): 3–20.
Rinehart, L.F., Heckert, A.B., Lucas, S.G., Hunt, A.P. (2004): The sclerotic ring of the Late Triassic theropod dinosaur Coelophysis. New Mexico Geological Society Spring Meeting. 26: 64.
Rinehart, L.F., Lucas, S.G., Heckert, A.B., Spielmann, J.A., Celesky, M.D. (2009): The paleobiology of Coelophysis bauri (Cope) from the Upper Triassic (Apachean) Whitaker quarry, New Mexico, with detailed analysis of a single quarry block. New Mexico Museum of Natural History & Science, A Division of the Department of Cultural Affairs Bulletin. 45: 260.
Rinehart, L. F., Lucas, S. G., Hunt, A. P. (2007): Furculae in the Late Triassic theropod dinosaur Coelophysis bauri. Paläontologische Zeitschrift. 81 (2): 174–180.
Sander, P. M. (2013): An Evolutionary Cascade Model for Sauropod Dinosaur Gigantism – Overview, Update and Tests. PLoS ONE 8(10): e78573.
Sander, P. M., Clauss, M. (2008): Sauropod gigantism. Science 322: 200–201.
Sander, P. M, Christian, A., Clauss, M., Fechner, R., Gee, C. T., et al. (2011): Biology of the sauropod dinosaurs: the evolution of gigantism. Biological Reviews 86: 117–155.
Schmitt, A. (2023): Großartige Giganten. Den letzten Geheimnissen der Dinosaurier auf der Spur. Dtv Verlagsgesellschaft
Schwartz, H. L., Gillette, D. D. (1994): Geology and taphonomy of the Coelophysis quarry, Upper Triassic Chinle Formation, Ghost Ranch, New Mexico. Journal of Paleontology. 68 (5): 1118–1130.
Schweitzer, M. H., Schroeter, E. R., Czajka, C. D. (2021): Dinosaurs How We KNow What We Know. CRC Press
Sellers, W. I., Margetts, L., Coria, R. A., Manning, P. L. (2013): March of the Titans: The Locomotor Capabilities of Sauropod Dinosaurs. PLoS ONE 8(10): e78733.
Sereno, P. C. (1999): The Evolution of Dinosaurs. Science 284(5423): 2137–2147
Sereno, P.C. (2010): Taxonomy, cranial morphology, and relationships of parrot-beaked dinosaurs (Ceratopsia:Psittacosaurus). In Ryan, Michael J., Chinnery-Allgeier, Brenda J., Eberth, David A. (eds.). New Perspectives on Horned Dinosaurs: The Royal Tyrrell Museum Ceratopsian Symposium. Bloomington and Indianapolis: Indiana University Press. pp. 21–58.
Sereno, P. C., Arcucci, A. B. (1994): Dinosaurian precursors from the Middle Triassic of Argentina: Marasuchus lilloensis, gen. nov. Journal of Vertebrate Paleontology. 14 (1): 53–73.
Sereno, P.C., Forster, C.A., Rogers, R.R., Moneta, A.M. (1993): Primitive dinosaur skeleton from Argentina and the early evolution of the Dinosauria. Nature. 361 (6407): 64–66.
Sereno, P. C., Martínez, Ricardo N., Alcober, Oscar A. (2013): Osteology of Eoraptor lunensis (Dinosauria, Sauropodomorpha). Basal sauropodomorphs and the vertebrate fossil record of the Ischigualasto Formation (Late Triassic: Carnian-Norian) of Argentina. Journal of Vertebrate Paleontology Memoir. 12: 83–179.
Sereno, P.C., Novas, F.E. (1992): The complete skull and skeleton of an early dinosaur. Science. 258 (5085): 1137–1140.
Sereno, P.C., Novas, F.E., Arcucci, A.B., Yu, C. (1988): New evidence on dinosaur and mammal origins from the Ischigualasto Formation (Upper Triassic, Argentina). Journal of Vertebrate Paleontology. 8 (3, supplement): 26A
Sterling J. Nesbitt, P. M. Barrett, Werning, S., et al (2012): The Oldest Dinosaur? A Middle Triassic Dinosauriform from Tanzania. Biology Letters. 9(1)
Stevens, K. A. (2013): The Articulation of Sauropod Necks: Methodology and Mythology. PLoS ONE 8(10): e78572.
Tang, F. et al. (2001): Biostratigraphy and palaeoenvironment of the dinosaur-bearing sediments in Lower Cretaceous of Mazongshan area, Gansu Province, China. Cretaceous Research. 22 (1): 115–129.
Taylor, M. P., Wedel, M. J. (2013a): The Effect of Intervertebral Cartilage on Neutral Posture and Range of Motion in the Necks of Sauropod Dinosaurs. PLoS ONE 8(10): e78214.
Taylor, M. P., Wedel, M. J. (2013b): Why sauropods had long necks, and why giraffes have short necks. PeerJ. 1: e36.
Thompson, R. S., Parish, J. C., Maidment, S. C. R., Barrett, P. M. (2011): Phylogeny of the ankylosaurian dinosaurs (Ornithischia: Thyreophora). Journal of Systematic Palaeontology 10(2): 301–312
Vinther, J., Nicholls, R., Kelly, Diane A. (2021): A cloacal opening in a non-avian dinosaur. Current Biology. 31 (4): R182–R183.
Vinther, J., Nicholls, R., Lautenschlager, S., Pittman, M., Kaye, T. G., Rayfield, E., Mayr, G., Cuthill, I. C. (2016): 3D Camouflage in an Ornithischian Dinosaur. Current Biology. 26 (18): 2456–2462.
Wedel, M. J., Taylor, M. P. (2013): Caudal Pneumaticity and Pneumatic Hiatuses in the Sauropod Dinosaurs Giraffatitan and Apatosaurus. PLoS ONE 8(10): e78213.
Weishampel, D. B., P. Dodson, and H. Osmolska, eds. (1990): The Dinosauria. 1st ed. Berkeley: University of California Press.
Xu, X., Forster, C. A., Clark, J. M., Mo, J. (2006): A basal ceratopsian with transitional features from the Late Jurassic of northwestern ChinaProc. R. Soc. B.2732135–2140
Yates, A. (2004): Anchisaurus polyzelus (Hitchcock): The smallest known sauropod dinosaur and the evolution of gigantism among sauropodomorph dinosaurs. Postilla. 230: 1–57.
Zanno, L. E. (2010a): Osteology of Falcarius utahensis (Dinosauria: Theropoda): characterizing the anatomy of basal therizinosaurs. Zoological Journal of the Linnean Society. 158 (1): 196–230.
Zanno, L. E. (2010b): A taxonomic and phylogenetic re-evaluation of Therizinosauria (Dinosauria: Maniraptora). Journal of Systematic Palaeontology. 8 (4): 503–543.
Zanno, L. E., Erickson, G. M. (2006): Ontogeny and life history of Falcarius utahensis, a primitive therizinosauroid from the Early Cretaceous of Utah. Journal of Vertebrate Paleontology. 26 (supp. 3): 143A.
