

Haben wir uns vielleicht geirrt? Sind Vögel doch keine Dinosaurier und hinter dieser Theorie steckt eine Verschwörung selbstgefälliger Paläontologen im Elfenbeinturm, die sich gegenseitig bestätigen und kritische Stimmen unterdrücken? Klingt das zu absurd um wahr zu sein? Nun, es gibt die „Birds are not Dinosaurs Bewegung“, die genau das behauptet. Ihr führender Vertreter ist Alan Feduccia, der seit Jahrzehnten vehement bestreitet, dass sich Vögel aus Dinosauriern entwickelt haben. Ist an dieser Verschwörung etwas dran? Bin ich selbst in diese Falle getappt oder gar Teil der Dino-Vogel-Verschwörung und wollte euch in den letzten sieben Episoden hinters Licht führen? Ihr werdet es in dieser Episode erfahren. Dieser Beitrag stützt sich zu einem großen Teil auf die Buchbesprechung von Darren Naish zu Feduccias neuestem Buch „Romancing the Birds and the Dinosaurs“ (Naish 2024)[1], welcher von der AG-Evolutionsbiologie ins Deutsche übersetzt wurde.[2]
Text als pdf
Wer ist Alan Feduccia?
John Alan Feduccia (Jahrgang 1943) ist ein Paläornithologe, der sich auf die Ursprünge und die Phylogenie der Vögel spezialisiert hat. Seit vier Jahrzehnten behauptet er, dass sich alle Welt über die Dinosaurier irre (Abb. 1). Feduccias Hauptthese lautet, dass Vögel sich nicht aus theropoden Dinosauriern entwickelt haben könnten, sondern ihren Ursprung in kletternden, vierfüßigen Nicht-Dinosaurier-Reptilien der Trias hätten.
Diese Meinung bringt er in Fachartikeln und Meinungsbeiträgen zum Ausdruck, vor allem aber in seinen Büchern „The Origin and Evolution of Birds“ (Feduccia 1996) und „Riddle of the Feathered Dragons“ (Feduccia 2012). Sein neuestes Buch ist „Romancing the Birds and Dinosaurs“ (Feduccia 2020), welches aus 23 Essays zum Stand der Dinosaurierforschung besteht, bzw. so wie es Feduccia sieht (Abb. 1).

Abb. 1: Alan Feduccia, der prominenteste Wissenschaftler, der in seinen Hauptwerken die Evolution der Vögel aus theropoden Dinosauriern abstreitet.
Zu Feduccias Leidwesen ist das „Vögel-sind-Theropoden“-Paradigma in der modernen Dinosaurierforschung fest verankert. Nicht nur, dass die Entstehung der Vögel als ein bedeutendes Ereignis in der Geschichte der Dinosaurier wahrgenommen wird; darüber hinaus wurden Elemente der Anatomie, des Verhaltens und der Biologie der Vögel in Dinosauriergruppen aller Art identifiziert. Anders ausgedrückt: Die Ansicht, dass Nichtvogel-Dinosaurier eng mit den Vögeln verwandt sind, ist Teil des Mainstream-Denkens. Und wir wissen ja: Mainstream-Denken ist immer schlecht und wenn ein Kritiker zu Wort kommt, muss doch was dran sein?
Mal im Ernst: der Konsens der Paläontologen, dass Vögel eine Untergruppe der Dinosaurier sind, liegt nicht daran, weil Paläontologen das sagen, oder weil sie sich miteinander verschworen haben und arme Dissidenten unterdrücken. Der Konsens ist Ausdruck der vorliegenden Daten, welche die Hypothese, dass Vögel Dinosaurier sind, am besten unterstützen, treffende Vorhersagen machen und der bisherigen Kritik standgehalten haben. Offene Forschungsfragen lassen sich ebenso wunderbar mit dem „Vögel sind Dinosaurier Modell“ erklären. Auf der anderen Seite haben alternative Hypothesen der Kritik nicht standgehalten.
Der Galileo-Mythos
Was Feduccia bzw. dessen Anhängern unterliegt, kann man sehr gut als „Galileo-Syndrom“ bzw. „Galileo-Fehlschluss“ bezeichnen. Das ist ein logischer Fehlschluss der besagt, dass ich Recht haben muss, wenn meine Ideen das Establishment dazu veranlassen, mich zu verunglimpfen oder zu bedrohen – „alle sagen, dass ich falsch liege, also habe ich Recht“ (Abb. 2).
Der Trugschluss bezieht sich auf die berühmte Verfolgung Galileo Galileis durch die römisch-katholische Kirche, weil er den Heliozentrismus gegen die damalige orthodoxe Bibelgläubigkeit verteidigte. Was die Befürworter dieses Trugschlusses nicht bedenken, ist, dass nicht alle Menschen, die den wissenschaftlichen Mainstream-Konsens in Frage stellen, Märtyrer oder Revolutionäre sind; in den meisten Fällen sind sie einfach nur Idioten: ich erinnere an Kreationisten, Flacherdler, Klimaleugner, Impfgegner usw. usf. Zugegeben: Feduccia ist (wahrscheinlich) kein Kreationist, dass aber ausgerechnet Kreationisten diejenigen sind, die sich am meisten auf ihn berufen ohne ihn aus dem Zusammenhang zu zitieren, ist schon verdächtig.
Nebenbei: Personen, die diesen Trick anwenden, um die Verfolgung durch die „große Wissenschaft“ zu behaupten, übersehen oft die Ironie in diesem Vergleich – es war die katholische Kirche, die Galileo zensierte, nicht das „wissenschaftliche Establishment“.
Der übliche Trugschluss beruht zum großen Teil darauf, dass die Weigerung der wissenschaftlichen Gemeinschaft, jeden Unsinn zu veröffentlichen oder sich mit jedem Spinner auseinanderzusetzen, als „Zensur“ missverstanden wird. Diese Masche kann viele Formen annehmen, aber in den meisten Fällen wird jemand, der damit für seine Ideen wirbt, seine vermeintliche Verfolgung hervorheben. Diese vermeintliche Verfolgung wird so lange aufgebauscht, bis ein Beobachter fast keine andere Wahl hat, als diese Ideen praktisch als Sympathiestimme zu akzeptieren. Genau solch eine Taktik gehen besonders Kreationisten ein, die ständig herumheulen, dass ihr Blödsinn von der Wissenschaft nicht ernst genommen wird. Dabei vergessen sie sehr oft, dass ihre Meinung eigentlich (und leider) gar nicht unterdrückt wird: schließlich können sie ihren Blödsinn ungehemmt im Internet verbreiten und die Leute manipulieren: und es lesen sich sicherlich mehr Leute blöde Social-Media-Posts oder schauen sich verrückte Prediger und Influencer auf YouTube an, als die mühselig zu lesenden wissenschaftlichen Publikationen zu Rate zu ziehen.
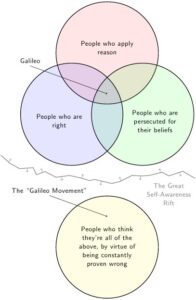
Abb. 2: Graphische Darstellung des Galileo-Fehlschusses
Eine zusätzliche Ironie ergibt sich, wenn wir bedenken, dass die Idee des Außenseiters, wenn sie es schafft, genügend Beweise zu sammeln, um die Mehrheit zu überzeugen, zum neuen Konsens wird. Die Vorstellung, dass Vögel Dinosaurier sind, war anfangs auch eine Minderheitenposition, setzte sich aber durch, weil sie viele empirische Daten am besten erklären konnte. Der Konsens der Paläontologen ist also Folge des Datenmaterials. Wie stellt aber Feduccia den Sachverhalt dar?
Er versucht vergeblich zu zeigen, dass die von Fachleuten vertretene Sicht auf die Dinosaurier chaotisch, oberflächlich und unreflektiert sei. Sie werde vom Streben nach Popularität und dem Festhalten an einem „kultähnlichen“ Kladismus, also der Kladistik, womit Feduccia ein enormes Problem zu haben scheint, bestimmt. Er vertritt die Ansicht, dass sich die Paläontologen größtenteils in allem irren. Besagte Paläontologen seien Teil einer Verschwörung, in der alle wesentlichen Arbeiten über vogelähnliche Dinosaurier und dinosaurierähnliche Vögel durch eine krasse Form der Bestätigungsverzerrung angetrieben würden: Feduccia unterstellt den Paläontologen, eine „theoriebeladene“, „monolithische“ Wissenschaft zu praktizieren und stellt wiederholt fest, dass Schlüsse gezogen würden, um „Kladogramme passend zu machen“, nur damit sie zu den vorgegebenen phylogenetischen Erzählungen passen.
Ohnehin vertritt Feduccia die Meinung, dass er und nur er sowie eine ausgewählte Gruppe von Kollegen die wahre Wissenschaft repräsentiere. Sie seien diejenigen mit der richtigen Methodik, jene, die einen aufrichtigen Skeptizismus praktizierten und die unabhängig von populären zeitgenössischen Trends arbeiteten. Sie allein könnten den Leser sicher durch das launische, oberflächliche, chaotische Gewirr an Behauptungen über Dinosaurier und archaische Vögel führen, die von den heutigen wissenschaftlich naiven, machthungrigen, jugendlichen Populisten aufgestellt würden.
Bestätigungsverzerrung Teil 1: Ich mach mir die (Vogel)welt, wie sie mir gefällt
Feduccia übersieht indes die Ironie, dass seine eigene Forschung tatsächlich von einer noch gezielteren Form der Bestätigungsverzerrung angetrieben wird als die, die er fälschlicherweise bei anderen identifiziert. Bestätigungsverzerrung ist übrigens eine Art der kognitiven Denkverzerrung, die dazu führt, dass Informationen einseitig verarbeitet und so interpretiert werden, so dass sie scheinbar die eigene (voreingenommene) Meinung bestätigen. Dabei werden Befunde, die nicht zur eigenen Meinung passen, außer Acht gelassen, für falsch oder unvollständig erklärt.
Wir haben in einer anderen Episode solch ein Beispiel vorgestellt:
Feduccia hält eine Verwandtschaft zwischen Vögeln und Dinosauriern für unmöglich (Feduccia 2002; Martin 2004; Feduccia et al. 2007). Gattungen wie Velociraptor, Deinonychus oder Oviraptor haben also unmöglich eine Nähe zu Vögeln. Nun ist es aber Tatsache, dass bei diesen Dinosauriern zweifelsohne Federn bekannt sind. Deswegen vertritt Feduccia seit neuestem eine völlig entgegengesetzte Position: Diese befiederten Raptoren sind überhaupt keine Dinosaurier, sondern sekundär flugunfähige Vögel … aber keine Dinosaurier. Es sei erwähnt: das einzige zusätzliche Merkmal, was bei diesen Gruppen neu dazu gekommen ist, sind Federn. Waren sie also vorher bloß Dinosaurier und nicht ansatzweise Vögel, sind sie nun bloße Vögel, aber nicht ansatzweise Dinosaurier. Es ist schwierig, dies ernst zu nehmen, wenn man bedenkt, dass nicht-aviale Maniraptoren offensichtliche Affinitäten zu nicht-avialen Coelurosauriern haben, die wiederum Affinitäten zu nicht-coelurosaurischen Theropoden haben, und so weiter und so weiter (Abb. 3). Hier haben wir einen wunderschönen Beleg für Feduccias Zurechtrücken von Fakten, um seinen Glauben, dass Vögel keine Dinosaurier sind zurechtzubiegen. Wenn das keine Bestätigungsverzerrung ist. Ich mache mir die Vogelwelt, wie sie mir gefällt.
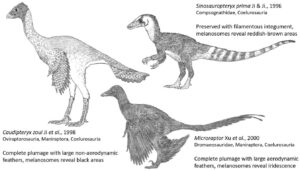
Abb. 3: Einige wichtige Theropoden-Taxa, die in „Romancing the Birds and Dinosaurs“ eingehend diskutiert werden. Sie alle zählen zur Theropoden-Gruppe der Coelurosaurier. Caudipteryx und Microraptor gehören zur Coelurosaurier-Untergruppe der Maniraptoren. Bild: Darren NAISH.
Die Idee, dass Maniraptoren wie Velociraptor die flugunfähigen Nachfahren von Vögeln sein könnten, stammt übrigens ursprünglich von Gregory Paul, der sie in seinem 2002 erschienenen Buch Dinosaurier der Lüfte am überzeugendsten darlegte (Paul 2002). Der entscheidende Unterschied: Gregory Paul hält sowohl die Raptoren als auch die Vögel für echte Dinosaurier. Feduccia hat somit eine verzerrte Hypothese von Paul übernommen, indem er die Dinosaurierverwandtschaft komplett ausklammert. Ironisch ist dies zudem, wenn klar wird, dass in der Vergangenheit Alan Feduccia gegenüber Gregory Paul nur Verachtung übrighatte.
Die meisten nachfolgenden kladistischen Analysen unterstützen Pauls Hypothese übrigens nicht. Beispielsweise wurde 2007 eine neue Gattung der Dromeosaurier beschrieben, Mahakala omnogovae (Turner et al. 2007, Abb. 4) welcher als ursprünglichster Dromeosaurier gilt – bislang galt als der basalste Dromeosaurier der flugfähige Microraptor (Hwang et al. 2002). Mahakala hatte kurze Arme und keine Fähigkeit zum Gleiten, weswegen die Autoren schlussfolgerten, dass sich der Flug nur bei den Avialae entwickelt hat, und der Vorfahr der Dromaeosauriden weder gleiten noch fliegen konnte. Im Jahr 2012 wurde jedoch in einer erweiterten und überarbeiteten Studie, die die jüngsten Dromaeosauridenfunde einbezog, der flugfähige Xiaotingia (Zhang et al. 2008, Xu et al. 2011, Abb. 5) als ursprünglichstes Mitglied der Dromaeosauridae-Klade identifiziert (Senter et al. 2012), was darauf hindeutet, dass die frühesten Mitglieder der Klade flugfähig gewesen sein könnten. (Einige ordnen Xiaotingia jedoch in die Anchiornithidae und mit diesen innerhalb der Avialae ein, siehe Foth & Rauhut 2017; andere ordnen Xiaotingia in die Nähe der Scansoriopterygidae innerhalb Paraves oder gar innerhalb der Avialae; siehe Lefevre et al. 2017; Eine Studie aus dem Jahr 2017 ergab, dass Mahakala ein Mitglied der rätselhaften, basalen Unterfamilie Halszkaraptorinae innerhalb der Dromeosauridae ist, siehe Cau et al. 2017). Ob Dromeosaurier bzw. alle Maniraptoren tatsächlich flugfähige Vorfahren hatten wird die zukünftige Forschung zeigen. Wichtig ist nur: Alle Untersuchungen belegen: Vögel sind Dinosaurier und die Maniraptoren ebenso.
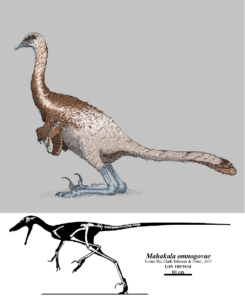
Abb. 4: Mahakala

Abb. 5: Xiaotingia
Bestätigungsverzerrung Nr. 2: Archeoraptor, Protoavis und Longisquama
In seinen Büchern erwähnt Feduccia regelmäßig den berüchtigten Archaeoraptor (Abb. 6) und beschuldigt damit die Paläontologen Fälschungen zu fördern, um das Paradigma der Vögel als Dinosaurier zu festigen, was nun Kreationisten in die Hände spielen. Worum geht es?
1999 machte die National Geographic Schlagzeilen, als ein „neuer Dinosaurier“ entdeckt wurde, der ein „fehlendes Bindeglied“ zwischen Vögeln und theropoden Dinosauriern darstellen soll: Archaeoraptor. Wenig später stellte sich heraus, dass es sich bei diesem Fossil um eine Fälschung handele und schon vor der Veröffentlichung gab es starke Zweifel an der Echtheit des Fossils. „National Geographic“ ist zwar seriöser als die „Bild“, jedoch ist es kein wissenschaftliches Fachjournal mit peer review – also nicht der richtige Ort für eine wissenschaftliche Erstbeschreibung. Studien fanden heraus, dass Archaeoraptor aus verschiedenen Fossilien zusammengesetzt wurde: der Oberkörper aus dem fossilen Vogel Yanornis und der Schwanz dem geflügelten Microraptor. Die Beine und Füße gehören zu einem unbekannten Tier (Zhou et al. 2002, Rowe et al. 2001).
Welche Auswirkungen hatte dieser peinliche Fehler auf Studien über archaische Vögel und ihrer Dinosaurier-Verwandten im Allgemeinen? Um es kurz zu machen, gar keine. Die einzigen, die diese falsche Fährte legen, sind die Kreationisten. Und natürlich Alan Feduccia.
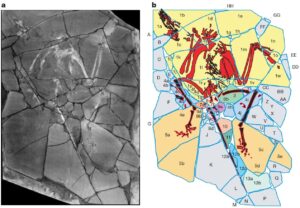
Abb. 6: In populären Medien wie dem National-Geographic-Magazin wurde der „Archaeoraptor“ als bedeutsames Mosaiksteinchen im Verständnis der Dinosaurier- und Vogelevolution gepriesen. ROWE et al. (2001) konnten jedoch nachweisen, dass es sich um eine aus verschiedenen Tierarten zusammengefügte Chimäre handelte, um ein gefälschtes Fossil also. Welche Auswirkungen hatte dieser peinliche Fehler auf Studien über archaische Vögel und ihrer Dinosaurier-Verwandten im Allgemeinen? Um es kurz zu machen, gar keine. Die einzigen, die diese falsche Fährte legen, sind die Kreationisten. Und natürlich Alan Feduccia. Bild: Rowe et al. (2001).
Ein Fossilfund könnte aber Feduccia vielleicht doch recht geben: Protoavis (Abb. 7), der erste Vogel, diesen erwähnt er in einem seiner älteren Bücher (Feduccia 1999). Es handelt sich hierbei um Skelettfunde aus der Obertrias, die in Texas gefunden wurden. Beschrieben wurden sie von Sankar Chatterjee, der diesen Fund als primitive Vogelart beschrieben habe (Chatterjee 1991, 1995, 1997). Würde dies zutreffen, hätte dies den Ursprung der Vögel um weitere 60 bis 75 Mio. Jahre zurückversetzt. Obwohl Protoavis damit wesentlich älter als Archaeopteryx ist, soll sein Skelettaufbau dennoch wesentlich vogelähnlicher sein. Diese Beschreibung von Protoavis setzt jedoch voraus, dass Protoavis korrekt als Vogel interpretiert wurde. Viele Paläontologen bezweifeln, dass es sich bei Protoavis um einen Vogel handelt oder dass alle ihm zugeordneten Überreste sogar von einer einzigen Art stammen, und zwar aufgrund der Umstände seiner Entdeckung und der nicht überzeugenden vogelartigen Synapomorphien in seinem fragmentarischen Material (Ostrom 1987, 1991, 1996, Nesbitt et al 2007). Als die Fossilien in Texas in den Sedimentschichten eines triassischen Flussdeltas gefunden wurden, handelte es sich um ein Sammelsurium disartikulierter Knochen, die möglicherweise auf ein Massensterben nach einer Sturzflut zurückzuführen sind (Chatterjee 1987).

Abb. 7: Protoavis
Zum anderen sind die Funde von Protoavis zu fragmentarisch, was eine genaue Zuordnung fragwürdig macht (Chiappe 1995, Welman 1995, Zhou 2004). Chatterjee war davon überzeugt, dass die Fragmente von zwei Individuen, einem jungen und einem alten Exemplar der gleichen Art gehörten. Es wurden jedoch nur wenige Teile gefunden, vor allem ein Schädel und einige Gliedmaßen, die zudem in ihren Proportionen zueinander nicht gut übereinstimmen, was viele zu der Annahme veranlasst hat, dass das Protoavis-Fossil eine Chimäre ist, also eine Mischung aus zwei oder mehr verschiedenen Arten.
Feduccia behauptet in der zweiten Auflage seines 1999 erschienenen Buchs „The Origin and Evolution of Birds“ (Feduccia 1999), dass Protoavis, sollte es sich als gültig erweisen, das Ende der theropoden Abstammung der Vögel bedeutet. Interessant ist hier anzumerken, dass selbst Chatterjee seinen vermeintlichen Vogel zur Verteidigung des Dinosaurier-Ursprungs der Vögel verwendet hat (Chatterjee 1997)! Er sah lediglich den Ursprung der Vögel von den Dinosauriern schon in der Trias. Somit widerspricht sogar Protoavis dem Argument von Alan Feduccia.
Ein anderer Favorit von Feduccia ist Longisquama, eine Gattung diapsider Reptilien, deren Fossilien 1970 in Sedimentgesteinen der Trias in Kirgisistan entdeckt wurden (Abb. 8). Die Funde basieren auf ein 15 cm langes Teilskelett mit fragmentiertem Schädel und Teilen des Rumpfskeletts. Besonders auffällig sind längliche blattartige Körperanhängsel, die von der Mittellinie des Rückens ausgehen. Fünf weitere Exemplare dieser Körperanhängsel ohne Zusammenhang zu Skelettfunden sind bisher beschrieben worden. Die taxonomische Einordnung dieser Gattung ist aufgrund der fragmentarischen Überlieferungen nicht ganz sicher. Einige ordnen Longisquama als Archosaurier, Archisauriformes, Lepidosauromorphen oder gar als frühen Diapsiden ein (Sharov 1970, Olshevsky 1991, Prum et al. 2001, Senter 2004, Buchwitz & Voigt 2012).
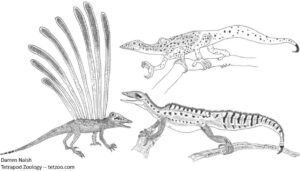
Abb. 8: Longisquama (links) mit auffällig langen, blattähnlichen Körperanhängseln und die Drepanosauriden (rechts) sind kleine vierfüßige, kletternde, diapside Reptilien aus der Trias. Sie sind weder Archosaurier noch deren nahe Verwandte. Es gibt daher keinen Grund anzunehmen, dass sie für die Abstammung der Vögel relevant seien. Feduccia und dessen Kollegen haben jedoch wiederholt auf die Möglichkeit verwiesen, dass diese Tiere näher mit den Vögeln verwandt seien als die (Nichtvogel-) Theropoden-Dinosaurier. Bild: Darren Naish.
Die Fragen zur Klassifizierung von Longisquama stehen auch im Zusammenhang mit dem Ursprung der Vögel. Feduccia behauptet, dass die verlängerten Körperanhängsel am Rücken von Longisquama für einen Gleitflug gedient haben könnten. Diese Annahme basiert auf einer frühen, mittlerweile nicht mehr zeitgemäßen Rekonstruktion aus den 1980er Jahren, welche davon ausgeht, dass es sich bei den Strukturen um lange, modifizierte Schuppen handelte, die paarweise an den Seitenwänden des Körpers angebracht waren, ähnlich wie gepaarte Gleitmembranen (Haubold & Buffetaut 1987), ähnlich wie bei den in Südostasien lebenden Gleitagamen der Gattung Draco (Abb. 9) oder beim ausgestorbenen triassischen Kuehneosaurus latus (Abb. 10) (Evans 2009, Pritchard & Nesbitt 2017, Colbert 1966, Stein et al. 2008).

Abb. 9: Gleitagamen der Gattung Draco.

Abb. 10: Kuhenosaurus
Eine Arbeitsgruppe um Feduccia (Jones et al. 2000, 2001) interpretierte diese Körperanhänge jedoch nicht als Schuppen, sondern als Federn. Die Mittelachse der Körperanhänge entspreche dem Federschaft und die Abzweigungen von der Mittelachse als Federfahne, wobei die einzelnen Strahlen der Fahne nur am Rand verwachsen seien. Weiterhin erinnere die Basis an den Federkiel moderner Vogelfedern. Die Deutung dieser Körperanhängsel als Federn hält ein Großteil der Paläontologen für eine Fehlinterpretation der Gewebsabdrücke im Gestein, die widersprechende Befunde nicht berücksichtigt (Prum 2001, 2002, Reisz & Sues 2000, Unwin & Benton 2001). So seien zum Beispiel nirgends Häkchen (Rami) nachzuweisen, die bei Kontur- und Schwungfedern die einzelnen Strahlen miteinander verbinden (so dass eine kontinuierliche Federfahne vorliegt). Randlich fusionierte Vogelfedern seien in der rezenten Vogelwelt ein sehr seltenes Phänomen und im vorliegenden Fall eine weit hergeholte Erklärung. Die als „Strahlen“ gedeuteten Strukturen der Anhängsel hätten z. T. keine Ähnlichkeit mit denen von Vogelfedern und seien an vielen Stellen nicht nachzuweisen. Am ehesten dürfte es sich also um verlängerte Rückenschuppen handeln.
Feduccias Hypothese, dass Vögel aus einer mysteriösen Gruppe triassischer Reptilien abstammen fußt also auf sehr dürftigen Funden und es ist bemerkenswert, dass er offensichtliche Gegenstimmen bewusst ignoriert.
Kladistik
Zudem zeigen alle phylogenetischen Untersuchungen, dass Vögel nun mal Dinosaurier sind. Was hat Feduccia dem entgegenzusetzen? Die Antwort: er lehnt per se die phylogenetischen Untersuchungen ab.
Sein Ziel ist es zu zeigen, dass das gesamte Feld chaotisch und unsicher sei und dass die taxonomischen Gruppen künstlich und schlecht definiert seien. Feduccia meint auch, die Homologie-Annahmen seien aufgrund falscher Merkmals-Gewichtungen nicht gerechtfertigt, die Taxon-Stichproben zu klein usw. Es gibt aber ausreichend Belege dafür (Smith et al. 2015), dass die Kladistik wohlfundiert ist und Zusatzwissen aus fast allen Biologiedisziplinen für die Plausibilitätsabwägung nutzt. Zahlreiche unabhängige Befunde aus Fachrichtungen wie Paläontologie, Physiologie, Histologie, Entwicklungsbiologie, Verhaltensbiologie etc. fügen sich ins Bild der Theropoden-Abstammung.
Ein philosophisches Problem, das bei Feduccia immer wieder auftaucht, ist sein Beharren darauf, dass die Gruppenzugehörigkeit in der Phylogenetik anhand einiger weniger, offensichtlicher, anatomischer Merkmale nachgewiesen werden sollte. Evolutionäre Gruppen sollen also typologisch bestimmt werden.
Bei Typologien (Abb. 11) werden einige wenige „typische“ Merkmale herangezogen, durch die eine bestimmte Gruppe konstituiert wird. Entscheidend ist hier weniger die tatsächliche Verwandtschaft, sondern eine phänotypische bzw. phänomenologische Klassifizierung nach analogen Eigenschaften, die zwar ein ähnliches Erscheinungsbild haben, aber nicht unbedingt verwandt sein müssen. So arbeitet aber die Klassifikation der Lebewesen in der biologischen Taxonomie nicht. Die Taxonomie und somit auch die Kladistik ist hierarchisch aufgebaut und bildet homologe Evolutionsprozesse ab. Dabei wird nach Ursprung, Entstehung oder Verwandtschaft klassifiziert und nicht nach oberflächlichen Ähnlichkeiten. PS: ein Beispiel für eine typologische Klassifikation, also eine, bei der evolutionäre Zusammenhänge keine tatsächliche Rolle spielen, ist die Einteilung des Menschen in irgendwelche Rassen.

Abb. 11: Unterschiede zwischen Typologie und Taxonomie
Organismen sind komplexe Einheiten – und wenn die Phylogenetik uns eines lehrt, dann, dass eine große Menge an Daten analysiert werden muss, wenn wir eine Phylogenie ermitteln wollen, die den wahren Verwandtschaftsverhältnissen möglichst nahekommt.
Die Identifikation von zwei oder drei „Schlüsselmerkmalen“ ist vollkommen unzureichend. Es ist erschreckend naiv anzunehmen, es sei gerechtfertigt, ein oberflächlich dinosaurierähnliches Reptil aufgrund des Besitzes einer langen Schnauze und spitzer Zähne als Dinosaurier zu charakterisieren, angesichts dutzender anatomischer Details, die ihm einen ganz anderen Platz im Kladogramm zuweisen.
Eine Reihe von Arten auf der Stammlinie der Krokodile, wie etwa der Poposauroide Effigia (Abb. 12 ganz oben im Diagramm), ähneln oberflächlich bestimmten Dinosauriern. Eine Fülle anatomischer Details belegt aber, dass sie nicht zur Stammlinie der Dinosaurier und Vögel gehören.
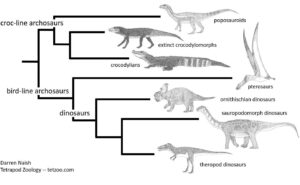
Abb. 12: Grob vereinfachte Darstellung der Archosaurier-Evolution, wie sie derzeit diskutiert wird. Eine Reihe von Arten auf der Stammlinie der Krokodile, wie etwa der Poposauroide Effigia ganz oben im Diagramm, ähneln oberflächlich bestimmten Dinosauriern. Eine Fülle anatomischer Details belegt aber, dass sie nicht zur Stammlinie der Vögel gehören und mit Dinosauriern nichts zu tun haben. Das hier dargestellte Kladogramm stützt sich auf eine Vielzahl von Studien, die eine enorme Anzahl anatomischer Beobachtungen und Entdeckungen beinhalten. Bild: Darren Naish.
Ein weiteres Beispiel für Strohmänner: Feduccia greift die phylogenetische Systematik aufgrund der Tatsache an, dass sich die Zuordnung einiger Taxa im Kladogramm immer wieder ändert. Feduccia will damit demonstrieren, dass die phylogenetische Standard-Systematik drastische Mängel aufweise. Ein Beispiel:
Lappentaucher und Seetaucher (Abb. 13) sind zwei Vogelfamilien, welche durch konvergente Evolution recht ähnlich aussehen. Beide Familien wurden ursprünglich in die Ordnung der Taucher zusammengefasst. Phylogenetische Untersuchungen haben jedoch gezeigt, dass beide Familien nicht näher miteinander verwandt sind. Die nächsten Verwandten der Lappentaucher sind die Flamingos, die Seetaucher werden in ein entferntes Verwandtschaftsverhältnis mit Albatrossen, Sturmvögeln oder Pinguinen gestellt (Jarvis et al. 2014). Heißt das aber jetzt, dass die Kladistik willkürlich und damit unbrauchbar ist?

Abb. 13 Es stimmt, dass Joel Cracraft einst Seetaucher (oben links), Lappentaucher wie den Haubentaucher (oben rechts) und die Hesperornithinen (unten) in der Klade der „Gaviomorphae“ zusammenfasste. Andere Autoren bildeten in jüngerer Zeit eine Klade aus Seetauchern und Haubentauchern. Beweisen solche Irrtümer, dass Versuche, die Stammesgeschichte der Vögel mit Hilfe der Kladistik zu rekonstruieren, zum Scheitern verurteilt und daher Zeitverschwendung sind? Nein, sie beweisen, dass die Wissenschaft fehleranfällig ist, aus Fehlern lernt und sich so allmählich „empor irrt“. Bilder: Seetaucher von John Picken from Chicago, USA, Gavia immer -Minocqua, Wisconsin, USA -swimming-8, Bild leicht beschnitten, CC BY 2.0. Die übrigen Bilder stammen von Darren Naish.
Das ist nicht das, was solche Beispiele zeigen. Sie zeigen, dass Phylogenetiker Fehler machen, wenn sie es versäumen, relevante Daten zu berücksichtigen oder diese Daten nicht bekannt sind. Dies ist kein fataler Fehler, sondern die unvermeidliche Folge wissenschaftlichen Fortschritts. Es ist unfair, dies als eine Art Rechtfertigung dafür zu benutzen, die Hände in den Schoß zu legen und mit der Wissenschaft aufzuhören. Es ist ja nicht so, dass Feduccias Bemühungen zur Rekonstruktion verwandtschaftlicher Beziehungen zwischen Organismen qualitativ bessere Resultate erbrächten. Falsche phylogenetische Hypothesen erstellte Feduccia in seiner eigenen Forschung zuhauf, so stellte er eine Verwandtschaft zwischen Schuhschnabel und den Störchen auf und eine zwischen Enten und Flamingos – beide Vorstellungen sind nicht mehr gültig (Feduccia 1977a, 1977b, 1978, Olson & Feduccia 1980a, 1980b).
Wissen wir was Theropoden sind?
Welche Absurditäten Feduccias Kritik an der Kladistik annehmen kann, zeigt folgendes Beispiel.
Kapitel sieben seines Buches „Romancing the Birds and Dinosaurs“ beginnt mit der Anekdote, in der Feduccia einen namentlich nicht genannten (!) Kollegen bittet, den Begriff Theropode zu definieren. Besagter Kollege soll die tiefschürfende Antwort gegeben haben: „Ein Theropode ist, was immer diese Leute über diesen sagen!“
Offenbar scheint Feduccia nicht zu interessieren, dass die Theropoden eine seit 140 Jahren anerkannte Untergruppe der Dinosaurier sind, die unzählige Male anhand apomorpher Merkmale definiert worden ist, in neuerer Zeit beispielsweise von Gauthier (1986), Sereno (1999), Rauhut (2003), Nesbitt et al. (2009), Cau (2018), eine wichtige Zusammenfassung liefern Holtz (2007, Kapitel 13) und Fastovsky & Weishampel (2021, Kapitel 6 bis 9).
So haben Theropoden ein verhältnismäßig großes Gehirn und ihre erste Zehe wurde stark verkleinert, sodass sie zum Laufen praktisch nutzlos war und die Füße möglichst schmal hielt, um besser Laufen zu können. Die hintere Hälfte des Schwanzes der frühen Theropoden war durch Knochenzapfen versteift, die halfen das Gleichgewicht bei schnellen Drehungen zu halten. Damit die Zähne beim Beutegreifen nicht zu sehr beschädigt werden, haben die Theropoden ein Intramandibulargelenk im Unterkiefer, also ein zusätzliches Gelenk zwischen dem zahnartigen Teil des Kiefers und den Knochen, die den Unterkiefer mit dem Oberkiefer verbinden (Abb. 14). Dieses zusätzliche Gelenk fungierte als Stoßdämpfer und entlastet die Zähne, wenn der Theropode in seine sich wehrende Beute zubeißt.
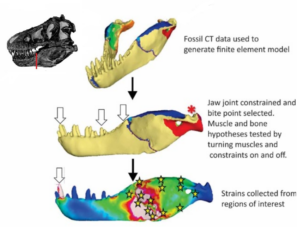
Abb. 14: Das Intramandibulargelenk bei Theropoden.
Der gemeinsame Vorfahre aller Dinosaurier hatte eine Greifhand, und bei den Theropoden wurden diese Hände auf das Töten spezialisiert. Die Klauen der Theropoden waren gekrümmt wie die Krallen von Raubvögeln. Weitere Apomorphien der Theropoden sind ihre hohlen Knochen und das Vorhandensein eines Gabelbeins. Betrachten wir uns nur die Maniraptora als Klade innerhalb der Theropoda. Zwei Besonderheiten seien hier erwähnt: ein halbmondförmiges Handgelenk, das aus der Verschmelzung mehrerer Handgelenksknochen besteht. Es dient als Hauptscharnier für die Bewegung des Handgelenks und ermöglicht die Handgelenke auszustrecken und Beutetiere mit einem schnellen Vor- und Zurückziehen zu greifen. Zufälligerweise ist genau die gleiche Bewegung Teil des Abwärtsflugschlags der Vögel. Im Gegensatz zu den meisten anderen Echsenbecken-Dinosauriern, deren Schambeine nach vorne zeigen, haben mehrere Gruppen von Maniraptoren ein nach hinten gerichtetes Schambein, ähnlich wie bei den Vögeln (Holtz & Osmolska 2004, Turner et al. 2007). Ich habe mich bewusst auf nur sehr wenige, für Laien leicht nachvollziehbare, Apomorphien begrenzt. Eine Studie von Andrea Cau (2018) listet 1431 Merkmale ausgehend von den Dinosauromorpha hin zu den modernen Vögeln, eingeteilt in 38 Kladen (!), auf, die nicht nur belegen, dass Vögel mitten im Stammbaum der Dinosaurier stehen, sondern Theropoden sehr gut definiert sind. Viele der aufgelisteten Merkmale sind sehr detailreich, und kladistisch gesehen sind Theropoden sehr gut definiert.
Jedoch bemerkt Feduccia, dass bestimmte Theropoden-Merkmale nicht bei allen Theropoden vorhanden seien, was ja ein Problem darstelle. Das ist entweder bemerkenswert naiv oder besonders trickreich: Die Evolution bedingt doch gerade, dass nicht alle Mitglieder einer Abstammungsgruppe Merkmale zeigen, die bei den frühesten Vertretern dieser Gruppe noch vorhanden waren. Erinnert sei an ein Beispiel aus einer anderen Tiergruppe: Schlangen. Sie zählen bekanntlich zu den Tetrapoden, also den vierfüßigen Landwirbeltieren. Aber Schlangen haben das Merkmal der vier Gliedmaßen verloren – dennoch sind sie Teil der Abstammungsgemeinschaft der Tetrapoden aus denen sie hervorgegangen sind. Warum sollte das bei Dinosauriern anders sein? Warum sollten bestimmte Merkmale bei kreidezeitlichen Theropoden gegenüber ihren triassischen Vorfahren nicht eine Veränderung erfahren haben?
Dollos Gesetz
Feduccia nimmt noch einen weiteren Taschenspielertrick aus der Mottenkiste. Er behauptet, dass alle nicht-maniraptorischen Theropoden „kurze, stummelartige Hände“ (S. 132, S. 163) hatten und ihre Vordergliedmaßen so reduziert waren, dass die Entwicklung langer Gliedmaßen, also Voraussetzung für die Entstehung der Vogelflügel, dem Dollo’schen Gesetz widersprechen würde.
Das Dollo’sche Gesetz besagt, dass eine gewisse Komplexität, einst verloren gegangen, keinesfalls in einem phylogenetisch jüngeren Organismus vollständig re-evolutioniert werden kann (Dollo 1893, siehe auch Gould 1970). Dazu mehrere Anmerkungen. Erstens zielt Dollos Gesetz auf Strukturen hin, die entweder völlig rudimentär (also praktisch nutzlos) oder ganz reduziert sind, etwa die Beine der Schlangen. Das ist bei den Armen der Theropoden nicht der Fall – die meisten Theropoden haben zwar relativ kurze, aber voll funktionelle Arme, die vermutlich für eine Vielzahl von Funktionen eingesetzt wurden.
Zum anderen hat sich Dollos ‚Gesetz‘ zwar als eine oft beobachtete Regel erwiesen, es ist aber nicht unumstößlich, wie zahlreiche Atavismen zeigen. Gerade die moderne Genetik (von der Dollo natürlich noch nichts wissen konnte) hat gezeigt, dass die genetischen ‚Programme‘ für viele reduzierte oder verlorene Organe oft immer noch vorhanden sind, aber nicht mehr abgerufen werden. Es gibt aber keinen Grund, weshalb sich so etwas nicht doch vielleicht manchmal wieder umkehren könnte (experimentell kann man zum Beispiel bei heutigen Vögeln durch Zugabe bestimmter Hormone in einem bestimmten Embryonalstadium die Ausbildung von Zahnanlagen auslösen, obwohl Vögel seit mindestens 80 Mio. Jahren zahnlos sind).
Tatsächlich aber haben schon jene Coelurosaurier, die keine Maniraptoren sind (dazu gehören die Ornithomimosaurier, Tyrannosauroiden und Compsognathiden), relativ lange, schlanke Hände, die bereits auf einen Trend zur Verlängerung der Vordergliedmaßen innerhalb dieser Gruppe hinweisen. Wenn man ehrlich ist, sehen sie so aus wie solche, von denen man annehmen würde, dass sie die Vorfahren der Maniraptoren sind (Abb. 15). Ja richtig gehört: sogar die ursprünglichen Tyrannosauroiden hatten ursprünglich relativ lange Gliedmaßen. Erwähnenswert ist hier Guanlong aus dem oberen Jura vor 160 Mio. Jahren (Xu et al. 2006, Abb. 16) – mit gerade mal 3m Länge und etwa 125 kg Lebendgewicht ein Zwerg zu seinem viel späteren, stummelarmigen Nachfolger Tyrannosaurus rex – aber er hatte relativ lange Arme mit drei Fingern.
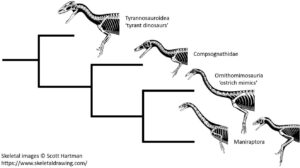
Abb. 15: Entgegen Feduccia zeigt sich bei den Theropoden, die auch die Maniraptoren umfassen, kein Trend zur Verkleinerung der Vordergliedmaßen. Wie das hier gezeigte Kladogramm andeutet, hatten sowohl Vertreter der Tyrannosauroidea (wie beispielsweise Guanlong) als auch die Ornithomimosaurier genauso wie die Maniraptoren relativ lange Arme im Vergleich zu Theropoden, die den Vögeln weniger ähneln. Ursprüngliche Maniraptoren (wie die Therizinosaurier) hatten dagegen proportional kürzere Arme als die vogelähnlicheren Maniraptoren. Bilder: © Scott HARTMAN, mit freundlicher Genehmigung.

Abb. 16: Guanlong
Scansoriopteryx
Es gibt eine Gruppe außergewöhnlicher Maniraptoren: die Scansoriopterygidae (Czerkas & Yuan 2002, Abb. 17). Sie sind faszinierende Tiere; sie vereinigen eine geringe Körpergröße mit einem kurzschnäuzigen Schädel, wahrscheinlichen Kletteranpassungen an den Hinterbeinen, mit langfingrigen Vorderbeinen, die (bei einigen Taxa, nicht bei allen) mit Flugmembranen ausgestattet sind. Ihre taxonomische Einordnung innerhalb der Maniraptora war (und ist es teilweise noch) nicht ganz sicher. Ursprünglich innerhalb der Avialae eingeordnet (Senter 2007, Zhang et al. 2008), stehen sie aktuelleren Untersuchungen außerhalb dieser und gelten als basale Paraves oder basale Pennaraptora (Agnolin & Novas 2011, 2013, Brusatte et al. 2014, Lefevre et al. 2014, Sorkin 2021, Pitmann & Xu 2020).
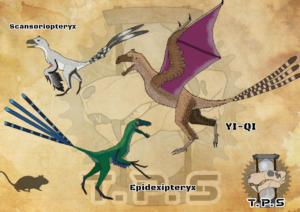
Abb. 17: Die Scansoriopterygiden sind wirklich faszinierende Tiere. Alle Vertreter sind klein, sie erreichen nur Sperlings- bis Taubengröße.
Sie sehen aus wie Vogelvorfahren, sofern man unbedingt will, dass sich die Vögel aus kleinen Baumkletterern entwickelt haben. So kann es nicht überraschen, dass sich Feduccia für sie interessiert. Das Problem für Feduccia ist nun, dass die Scansoriopterygiden zu den Theropoden aus der Gruppe der Maniraptoren zählen. Daher bedient sich Feduccia eines argumentativen Tricks (vgl. auch Czerkas & Feduccia 2014). Er besteht darin zu sagen, dass die Scansoriopterygiden „keine hervorstechenden Dinosauriermerkmale“ (S. 243) und „keine eindeutigen Theropoden-Merkmale“ (S. 247) besäßen. Zu den untypischen Theropoden-Merkmalen zählen ein nach vorne gerichtetes Schambein, das große Sitzbein, die weit auseinanderstehenden Darmbeine sowie die im Verhältnis zur Beinlänge relativ große Gesamtlänge der Arme. Zudem fehle den Scansoriopterygiden eine vollständig perforierte Hüftgelenkpfanne (Acetabulum), welches aber eine wichtige Apomorphie der Dinosaurier darstelle. Dieses Mosaik aus primitiven Skelettstrukturen und fortgeschrittenen Vogelmerkmalen nimmt Feduccia zum Anlass für eine gewagte Interpretation. Er bestreitet, dass es sich bei den Scansoriopterygiden um Dinosaurier handele und meint, dass sich die Vögel aus sehr ursprünglichen, Nicht-Dinosaurier-Archosauriern wie Scleromochlus entwickelten.
Auch hier konfrontiert uns Feduccia wieder mit seinen subjektiven Ansichten anstatt mit dem, was tatsächlich empirisch belegt ist. Zum einen zieht Feduccia für seine Einordnung nicht das reguläre Taxon der Dinosaurier heran, sondern wählt die typologische, an nur wenigen Schlüsselmerkmalen definierten Kategorie der Dinosaurier, die sich über einen idealtypischen Bauplan definiert. Dieses Problem hatten wir vorhin schon angesprochen. Weil die Scansoriopterygiden einige wenige evolutionäre Neuheiten aufweisen, die vom „Standardbauplan“ der ursprünglichen Dinosaurier abweichen, sind das nun keine Dinosaurier mehr. Weil Dinosaurier sich durch eine Hüftgelenkpfanne definieren, dürfen daraus abgeleitete, später evolvierte Arten davon nicht abweichen. Dieses typologische denken ist so sinnvoll wie die Behauptung, dass ein Merkmal der Vögel ihre Flugfähigkeit sei. Demnach dürften aber Strauße und Kiwis keine Vögel mehr sein – oder Schlangen keine Tetrapoden. Die Behauptung, dass sich die Scansoriopterygiden grundlegend von Theropoden und anderen Dinosauriern unterscheiden würden, ist schlicht falsch, da ihre Skelette mit Theropoden-Merkmalen förmlich übersät sind. Da aber auch sie eine eigenständige Evolution hatten, ist es nur folgerichtig anzunehmen, dass einige ihrer Merkmale, die typisch für basale Dinosaurier sind, dann auch verändert wurden.
Die erhebliche Subjektivität typologischer Betrachtungen ist der Grund, warum sie die Biologie heute praktisch einhellig ablehnt. Würde Feduccia die phylogenetische Systematik anwenden, erschienen die vermeintlich wesensmäßigen Unterschiede zwischen Scansoriopterygiden und Dinosauriern nur noch in Form abgestufter Ähnlichkeiten, zwischen denen etliche Theropoden vermitteln. Wie ordentliche Merkmalsanalysen gezeigt haben, gehörten die Scansoriopterygiden, wie alle Maniraptoren, den Coelurosauriern und damit den Dinosauriern an (Pittman & Xu 2020, Abb. 18).
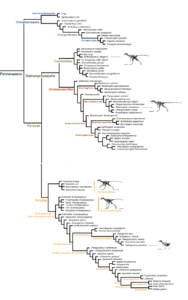
Abb. 18: Stammbaum der Pennaraptora nach Pittman & Xu (2020). Die Familie der Scansoriopterygidae ist hiernach Teil der Ovairaptorosauria.
Feduccia zerpflückt nicht nur das Theropoden-Taxon, sondern kreiert auch eine enorme Lücke innerhalb der ursprünglichen Avemetatarsalia: Die künstlich geschaffene Kluft zwischen triassischen Archosauriern wie Scleromochlus und vogelähnlichen Formen wie Scansoriopteryx müsste jetzt durch entsprechende Übergangsformen überbrückt werden (Abb. 19). Diese findet man unter den basalen Avemetatarsalia aber nicht, weil sie nur in der hoch diversifizierten Dinosaurier-Linie auftauchen. Die frühen Maniraptoren können diese Lücke auch nicht schließen, weil ausgerechnet diese (ungeachtet ihrer zahlreichen ursprünglichen Merkmale) von Feduccia zu sekundär flugunfähigen Vögeln, die keine Dinosaurier waren. umgedeutet werden! Phylogenetisch ist Feduccias Verwandtschaftshypothese damit äußerst unplausibel. Fatalerweise kommt hinzu, dass der von Feduccia favorisierte Verwandtschaftsbaum es erfordern würde, ein Maximum an Konvergenzen anzunehmen: Sämtliche Merkmale, die innerhalb der Ornithodira entstanden und sowohl bei Nichtvogel-Dinosauriern als auch bei Vögeln präsent sind, müssten parallel evolviert sein. Gewiss, bei nahen Verwandten ist immer mit Konvergenzen zu rechnen. Diese betreffen insbesondere die Reduktion und die Reversion einfacher bis mäßig komplexer Merkmale. Doch die Annahme, dass an der Basis der Avemetatarsalia-Stammgruppe sowie in der phylogenetisch weit entfernten Dinosaurier-Gruppe dutzende, wenn nicht hunderte anatomische Details parallel entstanden seien, ist völlig unglaubhaft. Wohlgemerkt, es geht nicht um die konvergente Entstehung oberflächlicher Strukturähnlichkeiten, die sich durch Funktionsgleichheit und Anpassung an gleiche Lebensräume erklärt. Solche analogen Merkmale wären beispielsweise die Flügel von Flugsauriern und Vögeln. Wir sprechen vielmehr von detaillierten Übereinstimmungen hierarchisch organisierter Merkmalskomplexe, die wir an anderen Stellen mehrmals aufgelistet haben.
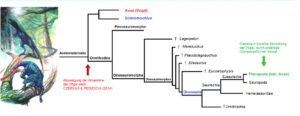
Abb. 19: Links: Zeichnerische Darstellung des Nicht-Dinosaurier-Archosauriers Scleromochlus, der noch relativ nah mit dem letzten gemeinsamen Vorfahren der Krokodile und Vögel verwandt ist. Rechts: Verwandtschaftsbaum der Avemetatarsalia mit der basalen Gattung Scleromochlus und der hochdiversen, tief ins Avemetatarsalia-Taxon eingeschachtelte Gruppe der Dinosaurier (inklusive der Theropoden). Entgegen der orthodoxen Auffassung stellen Czerkas & Feduccia (2014) die Vögel neben Scleromochlus an die Basis der Avemetatarsalia. Die Entwicklungslinie der Vögel hätte sich demnach schon sehr früh von den übrigen Archosauriern abgespalten (roter Pfeil); sie wären mit den Dinosauriern nicht näher verwandt. Ein Großteil der Vogelmerkmale wäre somit innerhalb der Dinosaurier-Linie konvergent entstanden. Grüner Pfeil: Kladistisch saubere Einordnung der Vögel, die auch durch unzählige Übergangsformen belegt ist. Quelle: Wikipedia | Urheber: Pavel.Ruha.CB | Lizenz CC BY-SA 3.0.
Doch keine Federn?
Eine weitere Debatte, in der Feduccia offenbar eine andere Sichtweise hat ist die Existenz von Keratin-Fasern bei Dinosauriern, die morphologisch und mikrostrukturell deutliche Ähnlichkeiten mit Federn aufweisen.
Feduccia ist bemüht, seine Leser glauben zu machen, dass es sich bei der flaumartigen Körperbedeckung gefiederter Dinosaurier, wie Sinosauropteryx (Currie & Chen 2001) in Wahrheit um Kollagenfasern der Haut handele (Abb. 20).
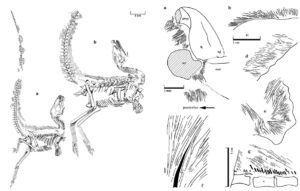
Abb. 20: In der Diskussion über den Ursprung der Vögel wird von einigen die Meinung vertreten, dass man dem Dinosaurier-“Flaum“ gegenüber sehr skeptisch sein sollte. Skepsis ist in dieser Frage tatsächlich angebracht. Aber regelrechte Verleumdungen auszusprechen, wie es Feduccia tat (Morell 1997, S. 38), war und ist ein Zeichen massiver Voreingenommenheit. Die Bilder zeigen zwei Sinosauropteryx-Exemplare mit fadenförmigen Integumentstrukturen [(a) NIGP 127586 und (b) NIGP 127587] sowie Nahaufnahmen dieser Filamente, die am gesamten Körper erhalten sind (und nicht nur an der dorsalen Mittellinie, wie Feduccia und Kollegen behaupten). Bilder: Currie & Chen (2001).
Aber selbst die großzügigste Auslegung der Daten zeigt, dass dies nicht zutrifft. Erstens, weil beiderlei Strukturen verschieden aussehen. Zweitens, weil der Dino-Flaum Merkmale zeigt, die darauf hindeuten, dass er aus Keratin und nicht aus Kollagen besteht. Und drittens, weil diejenigen, die andere davon überzeugen wollten, dass die Hautanhängsel aus Kollagen bestehen, sich sehr angestrengt und es vermasselt haben (man könnte auch sagen: geschummelt), um Kollagenfasern wie Dinosaurierflaum aussehen zu lassen.
Ich sage „geschummelt“, weil Feduccia anatomische Präparate abbildet, bei denen Kollagenfasern aus dem Knochen herausgelöst wurden, um Objekte zu erzeugen, die den Integument-Filamenten ähneln. Wesentlich für Feduccias Interpretation, dass es sich z. B. bei den Filamenten des Sinosauropteryx um Kollagenfasern handele, ist die Deutung, dass sich die Filamente „unterhalb der Haut“ (S. 185) bzw. innerhalb der „klar abgegrenzten Körperkonturen“ (S. 165) befänden (Abb. 21). Bei dem in Abb. 21 gezeigten Exemplar eines Sinosauropteryx (IVPP V12415) (das Bild zeigt einen Teil des Schwanzes) wurde die vermeintliche Haut- bzw. Körperkontur mit roten Pfeilen markiert. Tatsächlich aber handelt sich hier nicht um den Körperumriss, sondern um ein Präparations-Artefakt: Die Pfeile grenzen die Zone, in der die Gesteinsmatrix durch den Einsatz der Präparationswerkzeuge abblätterte, von dem Bereich ab, in dem die Matrix erhalten blieb. Dies ist bei all den betreffenden Exemplaren der Fall. Die meisten Beobachtungen, die zur Unterstützung der Kollagenfaser-Hypothese herangezogen werden, sind von diesem Kaliber.

Abb. 21: Wesentlich für Feduccias Interpretation, dass es sich z. B. bei den Filamenten des Sinosauropteryx um Kollagenfasern handele, ist die Deutung, dass sich die Filamente „unterhalb der Haut“ (S. 185) bzw. innerhalb der „klar abgegrenzten Körperkonturen“ (S. 165) befänden. Bei dem gezeigten Exemplar eines Sinosauropteryx (IVPP V12415) (das Bild zeigt einen Teil des Schwanzes) wurde die vermeintliche Haut- bzw. Körperkontur mit roten Pfeilen markiert. Tatsächlich aber handelt sich hier nicht um den Körperumriss, sondern um ein Präparations-Artefakt: Die Pfeile grenzen die Zone, in der die Gesteinsmatrix durch den Einsatz der Präparationswerkzeuge abblätterte, von dem Bereich ab, in dem die Matrix erhalten blieb. Dies ist bei all den betreffenden Exemplaren der Fall. Die meisten Beobachtungen, die zur Unterstützung der Kollagenfaser-Hypothese herangezogen werden, sind von diesem Kaliber. Bild: Darren Naish
Der einzige Fachmann, der Feduccias Deutung der Hautanhängsel teilte, war der inzwischen verstorbene Paläontologe Theagarten Lingham-Soliar. In den späten 1990er und frühen 2000er Jahren veröffentlichte Lingham-Soliar Arbeiten zur Beschaffenheit der Haut von Ichthyosauriern, wobei sein Hauptinteresse den komplexen, mehrschichtigen Kollagennetzen galt. Diese sind in die Haut eingebettet und bestehen aus sich überlappenden, gitterartigen Faserbündeln (Lingham-Soliar 2001, Lingham-Soliar & Wesley-Smith 2008, Abb. 22).
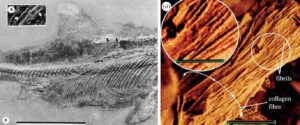
Abb. 22: Links: Ichthyosaurier-Hautfasern. Das Foto zeigt, dass die Fasern weit innerhalb des äußeren Körperumrisses liegen, das heißt tief in der Haut eingebettet sind. Rechts eine Nahaufnahme von Ichthyosaurier-Hautfasern, wie sie von Lingham-Soliar & Wesley-Smith (2008) zur Verfügung gestellt wurde. Man beachte, dass sich die zahlreichen winzigen Fibrillen nicht zu ihren Enden hin verjüngen, sondern zu dicken, seilartigen Strukturen vereinigen. Strukturell haben sie somit keine Ähnlichkeit mit dem Dinosaurier-Flaum. Bilder: Links: aus Lingham-Soliar (2001), Maßstab: 10 cm. Rechts: Lingham-Soliar & Wesley-Smith (2008), Maßstab: 2 Mikrometer; hineingezoomter Bereich: 1 Mikrometer.
Die Struktur dieser Faserbündel sieht nicht aus wie die Hautanhängsel der Dinos und Flugsaurier. Leider war Lingham-Soliar anderer Meinung; er veröffentlichte zwischen 2007 und 2015 Arbeiten und Bücher, in denen er wie Feduccia zu zeigen versuchte, dass es sich bei den Filamenten der Dinosaurier um verkannte Kollagenfasern handelt (Lingham-Soliar et al. 2007, Lingham-Soliar 2014, Feduccia et al. 2005). Wie also wurden Lingham-Soliars Argumente, abgesehen von Feduccia, rezipiert? Die primäre Reaktion war Ablehnung, denn gründlichere Untersuchungen ergaben, dass es sich um nicht-kollagene Fasern handelt, sondern um solche mit einzigartiger, für Haare und Federn typischer mikroskopischer Anatomie (Mayr 2010, Zhang et al. 2010, Godefroit et al. 2014, 2020, Mayr et al. 2016, Smithwick et al. 2017, Abb. 23, 24).

Abb. 23: Bilder oben: Bereiche mit außergewöhnlich gut erhaltenen haarartigen Hautanhängseln am Schwanz von Sinosauropteryx. Sie zeigen glatte, haarartige Filamente, die sich zu den Spitzen hin verjüngen, so wie dies für Keratin-Fasern (Haare, Federn) typisch ist, ohne Anzeichen von Wülsten, wie sie bei abgebautem Kollagen vorkämen. Bild unten links: Elektronenmikroskopische Aufnahme von Melanosomen, wie man sie in echten Federn des Vogels Confuciusornis fand. Unten rechts zum Vergleich Melanosomen, die in den Integument-Fasern von Sinosauropteryx gefunden wurden; die Ultrastrukturen sind identisch. Maßstab der elektronenoptischen Bilder: 2 Mikrometer. Bilder oben: aus Smithwick et al. (2017). Bilder unten: aus Zhang et al. (2010).

Abb. 24: Lingham-Soliar argumentierte, dass mehrere lange, gerade, nicht spitz zulaufende „Fasern“, die die Fossilien des Theropoden Sinosauropteryx umgeben, als Kollagenfasern identifiziert werden könnten. Die Merkmale, die er im Sinn hatte, entpuppten sich als Präparationsspuren von Werkzeugen, wie die hier gezeigten. Maßstab: 10 mm. Bilder: Smithwick et al. (2017).
Zhang et al. (2010) wiesen sogar fossile Melanosomen (Organellen von Pigmentzellen) in den Filamenten von Sinosauropteryx nach, die Federn und Haaren Farbe verleihen. Die Partikel entsprachen in Größe, Anordnung und Verteilung den Ultrastrukturen fossiler Melanosomen, die in echten Vogelfedern identifiziert wurden. Kollagenfasern enthalten dagegen keine Melanosomen. Und würden sie aus der Haut stammen, würde man eine homogene Verteilung erwarten, was nicht der Fall ist. Im Gegenteil, anhand der Verteilung ließ sich nachweisen, dass der Schwanz von Sinosauropteryx eine hell-dunkel-gestreifte Bänderung aufwies. Die Deutung als dermale Kollagenfasern ist daher abwegig.
Abschluss
An dieser Stelle machen wir Schluss. Feduccia zieht noch eine Reihe weiterer Argumente zu Rate: so die These der Warmblütigkeit der Dinosaurier oder die Homologie zwischen den Theropoden- und Vogelfingern. Aber diese wurden schon in den letzten Beiträgen behandelt und bedürfen keines weiteren Kommentars.
Alan Feduccia vertritt seit Jahrzehnten die Ansicht, dass nur er und seine Kollegen „die Vögel kennen“, während diejenigen, die das „Vögel-sind-Dinosaurier“-Paradigma vertreten, dies nicht tun. Das hat nie gestimmt, und es wird mit der Zeit immer offensichtlicher. Wie dem auch sei, wir könnten Feduccias Unterstellung problemlos spiegeln und ihm vorhalten, dass er mit Sicherheit von Nichtvogel-Dinosauriern keine Ahnung hat.
Literatur
Agnolín, F. L., Novas, F. E. (2011): Unenlagiid theropods: are they members of the Dromaeosauridae (Theropoda, Maniraptora)?. Anais da Academia Brasileira de Ciências. 83 (1): 117–162.
Agnolín, F. L., Novas, F. E. (2013): Avian ancestors. A review of the phylogenetic relationships of the theropods Unenlagiidae, Microraptoria, Anchiornis and Scansoriopterygidae. SpringerBriefs in Earth System Sciences. pp. 1–96.
Brusatte, S. L., Lloyd, G. T., Wang, S. C., Norell, M. A. (2014): Gradual assembly of avian body plan culminated in rapid rates of evolution across the dinosaur-bird transition. Current Biology. 24 (20): 2386–2392.
Buchwitz, M., Voigt, S. (2012): The dorsal appendages of the Triassic reptile Longisquama insignis: reconsideration of a controversial integument type. Paläontologische Zeitschrift. 86 (3): 313–331.
Cau, A. (2018): The assembly of the avian body plan: a 160-million-year long process. Bollettino della Società Paleontologica Italiana. 57 (1): 1–25.
Cau, A., Beyrand, V., Voeten, D. F. A. E., Fernandez, V., Tafforeau, P., Stein, K., Barsbold, R., Tsogtbaatar, K., Currie, P. J., Godefroit, P. (2017): Synchrotron scanning reveals amphibious ecomorphology in a new clade of bird-like dinosaurs. Nature. 552 (7685): 395–399.
Chatterjee, S. (1987): Skull of Protoavis and Early Evolution of Birds. Journal of Vertebrate Paleontology, 7(3)
Chatterjee, S. (1991): Cranial anatomy and relationships of a new Triassic bird from Texas. Philosophical Transactions of the Royal Society B: Biological Sciences, 332: 277-342.
Chatterjee, S (1995): The Triassic bird Protoavis. Archaeopteryx. 13: 15–31.
Chatterjee, S. (1997): The Rise of Birds: 225 Million Years of Evolution. Johns Hopkins University Press, Baltimore.
Chiappe, L. (1995): The first 85 million years of avian evolution. Nature. 391 (6555): 147–152.
Colbert, E. H. (1966): A gliding reptile from the Triassic of New Jersey. American Museum Novitates (3282): 1–23.
Currie, P. J., Chen, P.-J. (2001): Anatomy of Sinosauropteryx prima from Liaoning, northeastern China. Canadian Journal of Earth Sciences 38, 1705-1727.
Czerkas, S. A., Feducca, A. (2014): Jurassic archosaur is a non-dinosaurian bird. Journal of Ornithology, 155, 841–851.
Czerkas, S.A., Yuan, C. (2002): An arboreal maniraptoran from northeast China. S. 63-95 in Czerkas, S.J. (Ed.), Feathered Dinosaurs and the Origin of Flight. The Dinosaur Museum Journal 1. The Dinosaur Museum, Blanding, U.S.A
Dollo, L. (1893): Les lois de l’évolution. Bull. Soc. Belge Geol. Pal. Hydr. VII: 164–166.
Evans, S. E. (2009): An early kuehneosaurid reptile (Reptilia: Diapsida) from the Early Triassic of Poland. Paleontologica Polonica. 65: 145–178.
Fastovsky, D. E., Weishampel, D. B. (2021): Dinosaurs: A Concise Natural History (4th ed.). Cambridge university Press, Cambridge.
Feduccia, A. (1977a): The whalebill is a stork. Nature 266, 719-720.
Feduccia, A. (1977b): Hypothetical stages in the evolution of modern ducks and flamingos. Journal of Theoretical Biology 67, 715-721.
Feduccia, A. (1978): Presbyornis and the evolution of ducks and flamingos. American Scientist 66, 298-304.
Feduccia, A. (1996): The Origin and Evolution of Birds. Yale University Press, New Haven & London.
Feduccia, A. (1999): The Origin and Evolution of Birds (2nd ed.). Yale University Press, New Haven
Feduccia, A. (2002): Birds are dinosaurs: simple answer to a complex problem. Auk 119: 1187–1201.
Feduccia, A. (2012): Riddle of the Feathered Dragons: Hidden Birds of China. Yale University Press, New Haven & London.
Feduccia, F. (2020): Romancing the Birds and Dinosaurs: Forays in Postmodern Paleontology. BrownWalker Press, Irvine, Boca Raton.
Feduccia, A., Lingham-Soliar, T., Hinchliffe, J. R. (2005): Do feathered dinosaurs exist? Testing the hypothesis on neontological and paleontological evidence. Journal of Morphology. 266(2): 125–166.
Feduccia, A., Martin, L. D., Tarsitano, S. (2007): Archaeopteryx 2007: quo vadis? Auk 124: 373–380.
Foth, C., Rauhut, O.W.M. (2017): Re-evaluation of the Haarlem Archaeopteryx and the radiation of maniraptoran theropod dinosaurs. BMC Evolutionary Biology. 17 (1): 236.
Gauthier, J. A. (1986): Saurischian monophyly and the origin of birds. Memoirs of the California Academy of Sciences. 8: 1–55.
Godefroit, P., Sinista, S. M., Cinotta, A. et al. (2020): Integumentary structures in Kulindadromeus zabaikalicus, a basal neornithischian dinosaur from the Jurassic of Siberia. In FOTH, C. & RAUHUT, O. W. M. (Eds.) The Evolution of Feathers (pp. 48-65). Springer Nature, Berlin.
Godefroit, P., Sinista, S. M., Dhouailly, D. (2014): Response to Comment on ‚A Jurassic ornithischian dinosaur from Siberia with both feathers and scales‘. Science 346, 434.
Gould, S. J. (1970): Dollo on Dollo’s law: irreversibility and the status of evolutionary laws. Journal of the History of Biology. 3 (2): 189–212.
Haubold, H., Buffetaut, E. (1987): Une novelle interprétation de Longisquama insignis, reptile énigmatique du Trias supérieur d’Asie centrale [A new interpretation of Longisquama insignis, an enigmatic reptile from the Upper Triassic of Central Asia]. Comptes Rendus de l’Académie des Sciences de Paris. 305 (serie II): 65–70.
Holtz, T. R. (2007): Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages
Holtz, T.R., Osmólska, H. (2004): Saurischia. In Weishampel, Dodson and Osmólska (eds.), The Dinosauria, second edition. Berkeley: University of California Press.
Hwang, S.H., Norell, M.A., Ji, Q., Gao, K. (2002): New Specimens of Microraptor zhaoianus (Theropoda: Dromaeosauridae) from Northeastern China. American Museum Novitates (3381): 44pp.
Jarvis, E.D., et al. (2014): Whole-genome analyses resolve early branches in the tree of life of modern birds. Science. 346 (6215): 1320–1331.
Jones, T. D., Ruben, J. A., Maderson, P. F. A., Martin, L. D. (2001): Longisquama Fossil and Feather Morphology. Science 291(5510): 1901–1902
Jones, T. D., Ruben, J. A., Martin, L. D., Kurochkin, E. N., Feduccia, A., Maderson, P. F. A., Hillenius, W. J., Geist, N. R., Alifanov, V. (2000): Nonavian Feathers in a Late Triassic Archosaur. Science 288(5474): 2202–2205.
Lefèvre, U., Cau, A., Cincotta, A., et al. (2017): A new Jurassic theropod from China documents a transitional step in the macrostructure of feathers. The Science of Nature, 104: 74
Lefèvre, U., Hu, D., Escuillié, F. O., Dyke, G., Godefroit, P. (2014): A new long-tailed basal bird from the Lower Cretaceous of north-eastern China. Biological Journal of the Linnean Society. 113 (3): 790–804.
Lingham-Soliar, T. (2001): The ichthyosaur integument: skin fibers, a means for a strong, flexible and smooth skin. Lethaia 34, 287-302.
Lingham-Soliar T. (2014): Comment on „A Jurassic ornithischian dinosaur from Siberia with both feathers and scales“. Science 346(6208):434.
Lingham-Soliar T., Feduccia A., Wang X. (2007): A new Chinese specimen indicates that ‘protofeathers’ in the Early Cretaceous theropod dinosaur Sinosauropteryx are degraded collagen fibresProc. R. Soc. B.2741823–1829
Lingham-Soliar, T., Wesley-Smith, J. (2008): First investigation of the collagen D-band ultrastructure in fossilized vertebrate integument. Proceedings of the Royal Society B 275, 2207-2212.
Martin, L. D. (2004): A basal archosaurian origin for birds. Acta Zoologica Sinica 50: 978–990.
Mayr, G. (2010): Response to Lingham-Soliar: dinosaur protofeathers: pushing back the origin of feathers into the Middle Triassic? Journal of Ornithology 151, 523-524.
Mayr, G., Pitman, M., Saitta, E. T. et al. (2016): Structure and homology of Psittacosaurus tail bristles. Palaeontology 59, 793-802.
Morell, V. (1997): The origin of birds: the dinosaur debate. Audubon 99 (2), 36-45.
Naish, D. (2023): Alan Feduccia (2020): ‚Romancing the Birds and Dinosaurs‘. Deutsche Übersetzung: https://www.ag-evolutionsbiologie.net/html/2023/feduccia-romancing-birds-and-dinosaurs.html englisches Original: https://tetzoo.com/blog/2023/10/2/alan-feduccia-romancing-the-birds-and-dinosaurs
Nesbitt, S. J., Irmis, R. B., Parker, W. G. (2007): A critical re-evaluation of the Late Triassic dinosaur taxa of North America. Journal of Systematic Palaeontology. 5 (2): 209–243.
Nesbitt, S. J., Smith, N. D., Irmis, R. B. et al. (2009): A complete skeleton of a late Triassic Saurischian and the early evolution of dinosaurs. Science 326, 1530-1533.
Olshevsky, G. (1991): A Revision of the Parainfraclass Archosauria Cope, 1869, Excluding the Advanced Crocodylia. Mesozoic Meanderings, 2: 196 pp
Olson, S. L., Feduccia, A. (1980a). Relationships and evolution of flamingos (Aves: Phoenicopteridae). Smithsonian Contributions to Zoology 316, 1-73.
Olson, S. L., Feduccia, A. (1980b). Presbyornis and the origin of the Anseriformes (Aves: Charadriomorphae). Smithsonian Contributions to Paleobiology 323, 1-24.
Ostrom, J.H. (1987): Protoavis, a Triassic bird?. Archaeopteryx. 5: 113–114.
Ostrom, J.H. (1991): The bird in the bush. Nature. 353 (6341): 212.
Ostrom, J.H. (1996): The questionable validity of Protoavis. Archaeopteryx. 14: 39–42.
Paul, G.S. (2002): Dinosaurs of the Air: The Evolution and Loss of Flight in Dinosaurs and Birds. Baltimore: Johns Hopkins University Press. p. 257.
Pittman, M., Xu, X. (2020): Pennaraptoran Theropod Dinosaurs Past Progress and New Frontiers. Bulletin of the American Museum of Natural History. 440 (1): 1–355.
Pritchard, A. C., Nesbitt, S. J. (2017): A bird-like skull in a Triassic diapsid reptile increases heterogeneity of the morphological and phylogenetic radiation of Diapsida. Royal Society Open Science. 4 (10): 170499.
Prum, R. O. (2001): Longisquama Fossil and Feather Morphology. Science 291(5510): 1899–1900
Prum, R. O. (2002): Are current critiques of the theropod origin of birds science? Rebuttal to Feduccia. The Auk. 120 (2): 550–561.
Prum, R. O., Unwin, D.M., Benton, M.J. (2001): Response, Jones, T.D., Ruben, J.A., Maderson, P.F.A., Martin, L.D. (2001). Longisquama Fossil and Feather Morphology. Science. 291 (5510): 1899–1902.
Rauhut, O. W. M. (2003): The interrelationships and evolution of basal theropod dinosaurs. Special papers in paleontology 69, 1-213.
Reisz, R.R., Sues, H.-D. (2000): The „Feathers“ of Longisquama. Nature. 408 (6811): 428.
Rowe, T., Ketcham, R. A., Denison, C. et al. (2001): Forensic palaeontology: The Archaeoraptor Forgery. Nature 410, 539 – 540
Senter, P. (2004): Phylogeny of the Drepanosauridae (Reptilia: Diapsida). Journal of Systematic Palaeontology. 2 (3): 257–268.
Senter, P (2007): A new look at the phylogeny of Coelurosauria (Dinosauria: Theropoda). Journal of Systematic Palaeontology. 5 (4): 429–463.
Senter, P., Kirkland, J. I., Deblieux, D. D., Madsen, S., Toth, N. (2012): New Dromaeosaurids (Dinosauria: Theropoda) from the Lower Cretaceous of Utah, and the Evolution of the Dromaeosaurid Tail. PLOS ONE. 7 (5): e36790.
Sereno, P. C. (1999): The evolution of dinosaurs. Science. 284 (5423): 2137–2147.
Sharov, A.G. (1970): A peculiar reptile from the lower Triassic of Fergana. Paleontologicheskii Zhurnal (1): 127–130.
Smith, N. A., Chiappe, L. M., Clarke, J. A., et al. (2015): Rhetoric vs. reality: a commentary on “Bird origins anew” by A. Feduccia. The Auk: Ornithological Advances, 132, 467–480.
Smithwick, F. M., Mayr, G., Saitta, E. T. et al. (2017): On the purported presence of fossilized collagen fibres in an ichthyosaur and a theropod dinosaur. Palaeontology 60, 409-422.
Sorkin, B. (2021): Scansorial and aerial ability in Scansoriopterygidae and basal Oviraptorosauria. Historical Biology. 33 (12): 3202–3214.
Stein, K., Palmer, C., Gill, P. G., Benton, M. J. (2008): The aerodynamics of the British Late Triassic Kuehneosauridae. Palaeontology. 51 (4): 967–981.
Turner, A. H., Pol, D., Clarke, J. A., Erickson, G. M., Norell, M. (2007): A basal dromaeosaurid and size evolution preceding avian flight. Science. 317 (5843): 1378–1381.
Unwin, D. M., Benton, M. J. (2001): Longisquama fossil and feather morphology. Science 291, 1900-1901.
Welman, J (1995): Euparkeria and the origin of birds. South African Journal of Science. 91: 533–537
Xu X., Clark, J.M., Forster, C. A., Norell, M.A., Erickson, G.M., Eberth, D.A., Jia, C. & Zhao, Q. (2006): A basal tyrannosauroid dinosaur from the Late Jurassic of China. Nature. 439 (7077): 715–718.
Xu, X., You, H., Du, K. et al. (2011): An Archaeopteryx-like theropod from China and the origin of Avialae. Nature 475, 465–470.
Zhang, H., Wang, M., Liu, X. (2008): Constraints on the upper boundary age of the Tiaojishan Formation volcanic rocks in West Liaoning-North Hebei by LA-ICP-MS dating. Chinese Science Bulletin. 53 (22): 3574–3584.
Zhang, F., Kearns, S. L., Orr, P. J. et al. (2010): Fossilized melanosomes and the colour of Cretaceous dinosaurs and birds. Nature 463, 1075-1078.
Zhang, F., Zhou, Z., Xu, X., Wang, X., Sullivan, C. (2008): A bizarre Jurassic maniraptoran from China with elongate ribbon-like feathers. Nature. 455 (7216): 1105–1108.
Zhou, Z. (2004): The origin and early evolution of birds: discoveries, disputes, and perspectives from fossil evidence. Naturwissenschaften. 91 (10): 455–471.
Zhou, Z., Clarke, J. A., Zhang, F. (2002): Archaeoraptor’s better half. Nature 420: 285.
[1] https://tetzoo.com/blog/2023/10/2/alan-feduccia-romancing-the-birds-and-dinosaurs
[2] https://www.ag-evolutionsbiologie.net/html/2023/feduccia-romancing-birds-and-dinosaurs.html
