

Die Vogelwelt bietet immer wieder überraschende Einblicke in die Evolution. Eine besonders faszinierende Erkenntnis der modernen Molekularphylogenie ist die nahe Verwandtschaft von Vögeln, die auf den ersten Blick kaum Gemeinsamkeiten aufweisen: Falken, Seriemas, die ausgestorbenen Terrorvögel (Phorusrhacidae), Papageien und Singvögel. Diese scheinbar disparate Gruppe bildet die Klade der Australaves. Die überraschendste Erkenntnis ist hierbei, dass die Falken trotz ihrer Ähnlichkeiten mit Greifvögeln mit Papageien und Spatzen enger verwandt sind als mit Adlern und Bussarden. Falken und Greifvögel sind ein Paradebeispiel für konvergente Evolution.
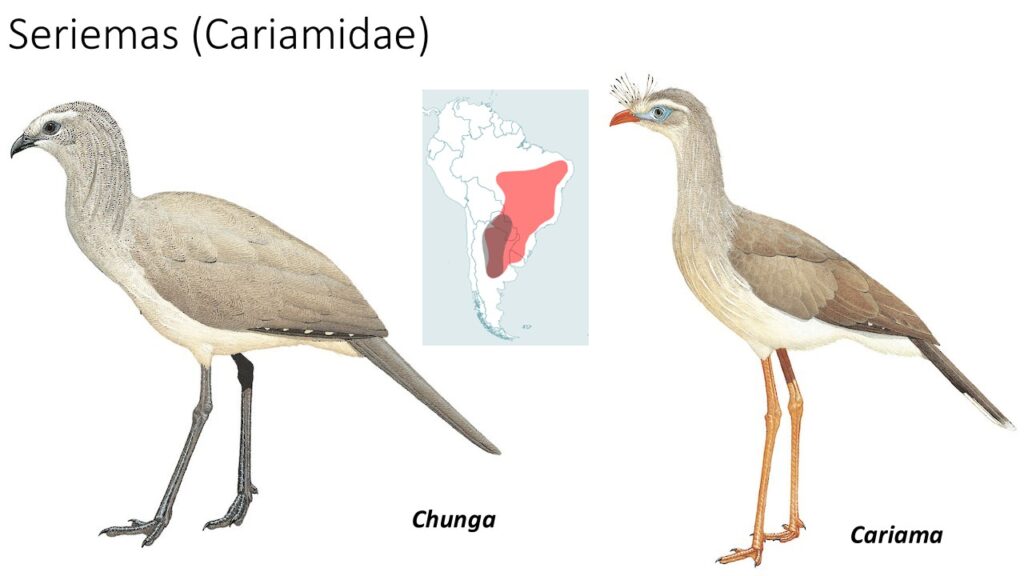
Seriemas (Cariamidae)
In der Ordnung der Cariamiformes gibt es nur eine Familie mit zwei Gattungen mit je einer Art: die Seriemas (Cariamidae). Es sind vorwiegend bodenlebende Vögel, die in trockenen Landschaften Südamerikas verbreitet sind. Mit einem durchschnittlichen Gewicht von 1,5 kg und einer Körperlänge von bis zu 90 cm ist die Rotfußseriema (Cariama cristata) etwas größer und schwerer als die Schwarzfußseriema (Chunga burmeisteri), der nur etwa 80 cm lang, und 1,2 kg schwer ist. Die langen und schlanken Beine haben drei kräftige nach vorn gerichtete Zehen und eine kleinere Hinterzehe (Anisodactylie). Mit ihrem kräftigen und gebogenen Schnabel ernähren sie sich vor allem von Insekten aber auch Nagetiere und Schlangen. Hin und wieder werden Früchte und Samen gefressen (Del Hoyo 2020, Del Hoyo et al. 1996, Winkler 2020a, Winkler et al. 2015).
Die taxonomische Einordnung der Seriemas war lange Zeit umstritten und sie wurden in die Verwandtschaft mit Kranichen, Sekretären, Trappen oder Kagus gestellt. Die Ähnlichkeiten zu diesen Vögeln beruhen aber auf konvergente Evolution und sie stellen tatsächlich die basalste Gruppe innerhalb der Australaves dar und die drei anderen Ordnungen (Falken, Papageien und Sperlingsvögel) werden als Eufalconimorphae zusammengefasst und den Seriemas gegenübergestellt (Cracraft et al. 2004, Hackett et al. 2008, Jarvis et al. 2014, Prum et al. 2015).
Seriemas haben eine lange Evolutionsgeschichte in Südamerika, und ihr ältester Vertreter, Noriegavis aus dem frühen bis mittleren Miozän von Patagonien, ist den heute lebenden Arten bereits sehr ähnlich. Allerdings stellen die Seriemas nur ein kleines Segment der früheren Vielfalt der Cariamiformes dar. Denn in diese Ordnung gehören eine Reihe ausgestorbener Vertreter, deren bekannteste Familie die Phorusrhacidae, die Terrorvögel, sind (Mayr & Noriega 2013. Alvarenga & Höfling 2003, Cenizo 2012, LaBarge et al. 2024, Mayr 2016).
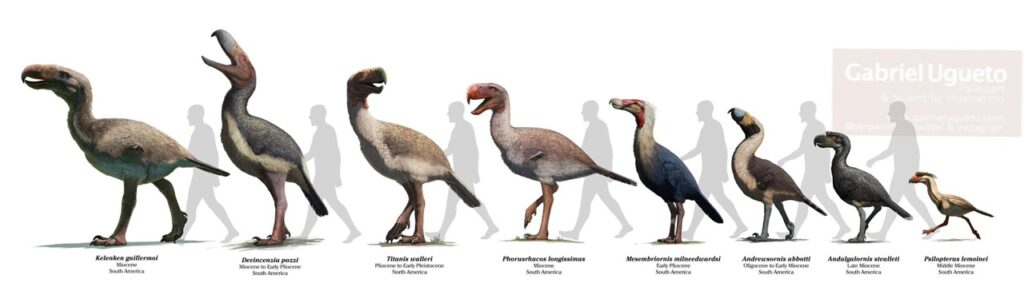
Terrorvögel
In Südamerika gab es diese charakteristischen flugunfähigen Vögel vom Paläozän bis zum Pliozän, und einige wenige Arten haben sogar bis ins späte Pleistozän überlebt. Terrorvögel zeichnen sich u. a. durch ihre massiven, raubvogelähnlichen Schnäbel und die Reduktion der Flügelknochen aus. Die meisten Arten haben lange Hinterbeine, was auf eine bodenbewohnende Lebensweise hindeutet. Einige frühere Gattungen könnten noch eine beschränkte Flugfähigkeit gehabt haben. Terrorvögel gehörten zu den wichtigsten terrestrischen Raubtieren im Känozoikum Südamerikas. Die Schädelmerkmale deuten darauf hin, dass sie entweder Beutetiere fraßen, die ganz geschluckt werden konnten, oder Beutetiere durch mehrere Schläge mit dem riesigen Schnabel töteten. Die meisten Fossilien stammen aus Argentinien, aber es gibt auch Funde aus Brasilien und Uruguay. Derzeit werden etwa 20 Arten unterschieden, die in die Unterfamilien Mesembriornithinae („Hermosiornithinae“), Psilopterinae, Patagornithinae, Phorusrhacinae und Physornithinae eingeordnet werden. Hinzu kommen einige weitere Gattungen, deren taxonomische Position nicht sicher ist (Mayr 2016, Alvarenga 2015, Alvarenga & Höfling 2003, Cenzio 2012, LaBarge et al. 2024, Mayr & Noriega 2013, Acosta Hospitaleche & Jones 2024, Degrange 2020, Degrange et al. 2015, 2025, Prevosti et al. 2013, Tarquini et al. 2022).
Zu den frühesten Vertretern gehört Paleopsilopterus aus dem Paläozän. Er gehört in die Unterfamilie Psilopterinae, wie die Gattung Psilopterus aus dem Ologozän und Miozän. Psilopterus war einer der kleinsten Terrorvögel, etwa so groß wie der Rotfußseriema. Ihre Flügelknochen waren weniger reduziert, als die anderer Terrorvögel. Außerdem war auch das Gabelbein gut entwickelt, was auf eine Flugfähigkeit hinweist.
Die Unterfamilien Patagornithinae und Phorusrhacinae haben eine lange Unterkiefersymphyse und einen langen Tarsometatarsus, wobei die Patagornithinae kleiner waren. Zu den Patagornithinae gehören Gattungen wie Patagornis, Andalgalornis und Andrewsornis. Zu den Phorusrhacinae zählen Gattungen wie Phorusracos und die recht großen Gattungen Devincenzia und Kelenken. Letzterer wurde bis zu 3m hoch und 100 kg schwer. Nach der Bildung der mittelamerikanischen Landbrücke verbreitete sich Terrorvogel, der bis zu 2m hohe Titanis, nach Nordamerika, wo es in pliozänen Ablagerungen, also vor 5 – 1.8 Mio. Jahren, in Texas und Florida gefunden wurde. In die Unterfamilie Mesembriornithinae gehören Mesembriornis („Hermosiornis„), Llallawavis und Procariama. Zu den Physornithinae gehören die Gattungen Physornis und Paraphysornis. Früher wurden sie zusammen mit Brontornis als Unterfamilie Brontornithinae zusammengefasst. Brontornis wird aber von den meisten nicht mehr als Terrorvogel eingeordnet, sondern gehört in die Verwandtschaft der Galloanserae.
Das Aussterben der Terrorvögel steht wahrscheinlich mit der Bildung der mittelamerikanischen Landbrücke und der einhergehenden Einwanderung nordamerikanischer Arten nach Südamerika im Zusammenhang. Die Terrorvögel könnten entweder von fleischfressenden Säugetieren gejagt worden sein oder mit ihnen um Beutetiere konkurriert haben. Die Hauptbeute der Terrorvögel, die südamerikanischen Huftiere, Notoungulata, starben zu einem ähnlichen Zeitpunkt aus. Die Tatsache, dass mindestens ein Terrorvogel-Taxon, Titanis, nach der Bildung der mittelamerikanischen Landbrücke nach Nordamerika eingewandert ist, stellt jedoch diese Vorstellungen in Frage. Alternativ wurde die Hypothese aufgestellt, dass der Wettbewerb mit Kondoren eine Rolle beim Aussterben der Terrorvögel gespielt hat, was eine brauchbare Hypothese wäre, wenn diese Vögel Aasfresser waren, aber auch das Vorkommen von Titanis in Nordamerika nicht erklären kann.
Gab es Terrorvögel außerhalb Amerikas?
Es gibt vermutliche Überreste von Terrorvögeln aus dem späten Eozän von Seymour-Island, die zur Antarktis gehört. Da es zu dieser Zeit eine Landverbindung zwischen der Antarktis und Südamerika gab, wäre das Vorkommen von Terrorvögeln in der Antarktis nicht unerwartet (Angst et al. 2013, Mayr 2002, 2005, 2016, Mayr & Richter 2011, Mayr et al. 2013, Peters 1987, Stidham & Wang 2017, Mourer-Chauviré et al. 2011).
Noch rätselhafter sind die Nachweise von mutmaßlichen Terrorvögeln in der Alten Welt. Bei diesen Fossilien gibt es Ähnlichkeiten in der Morphologie der Terrorvögel, z. B. im Bereich der Flügel und Brust, es gibt aber auch Unterschiede. Einer dieser Gattungen ist Strigogyps aus der Grube Messel, von dem drei Arten beschrieben sind. Es sind hühnergroße Vögel mit kräftigen Beinen und reduzierten Flügeln. Details des Tibiotarsus weisen darauf hin, dass Strigogyps eine gut entwickelte Zehenstreckmuskulatur hatte und seine Füße zur Manipulation von Nahrungsmitteln verwendete. Das sind Merkmale die dieses Taxon von den Terrorvögeln deutlich unterscheiden. Bei einem Exemplar wurden auch Pflanzenmaterial als Mageninhalt gefunden.
Eine andere Gattung ist Salmila (Salmilidae) aus der Grube Messel, der wahrscheinlich mit Strigoyps verwandt ist, aber kleiner als dieser und flugfähig ist (Mayr 2009). Strigogyps-ähnliche Vögel wurden auch aus dem Paläozän Chinas (Qianshanornis, Qianshanornithidae) und Oligozän Ägyptens beschrieben.
Ein anderer potentieller Terrorvogel aus dem Eozän der Schweiz ist Eleutheornis, der zuvor zu den Laufvögeln gerechnet wurde. Seine Funde sind aber zu fragmentarisch für eine genauere Zuordnung, aber seine Krallen ähneln Strigogyps (Angst et al. 2013, Mayr 2009a).
Lavocatavis aus dem Eozän Algeriens ist ein weiterer flugunfähiger Terrorvogel-Kandidat und wird durch einen einzigen Oberschenkelknochen repräsentiert. Es ist aber nicht auszuschließen, dass Lavocatavis eher mit Eremopezus, einem flugunfähigen Palaeognathae aus dem Eozän Nordafrikas, verwandt ist. Zusammenfassend lässt sich sagen, dass es keine eindeutigen Nachweise von Terrorvögeln aus der Alten Welt gibt. Darüber hinaus sind keine etablierten und weithin akzeptierten Ausbreitungswege über Land bekannt, die die Verbreitung der Terrorvögel zwischen Europa und Südamerika im frühen Paläogen ermöglicht hätten.
Europäische und nordamerikanische Cariamiformes
Ungeachtet der umstrittenen Anwesenheit von Terrorvögeln gab es in Europa während des frühen und mittleren Känozoikums unbestrittene Stammgruppen der Cariamiformes. Eine besonders gut vertretene Gruppe von ihnen sind die Idiornithidae (Mayr 2009, 2016, 2016a, 2017, 2022, Mayr et al. 2011, Cracraft 1968, Labarge et al. 2024).
Bis vor kurzem wurden diese Seriema-ähnlichen Vögel dem Taxon „Idiornis“ zugeordnet. Inzwischen hat man jedoch erkannt, dass die im späten 19. Jahrhundert als Dynamopterus beschriebenen Oberarmknochen zur selben Art gehören wie andere Knochen, die „Idiornis“ zugeordnet wurden. Da Dynamopterus nomenklatorisch Vorrang hat, wurden alle derzeit anerkannten „Idiornis„-Arten in dieses Taxon überführt.
Aus Frankreich wurden sechs Dynamopterus-Arten unterschieden und weitere Arten aus dem Eozän Deutschlands. Alle stimmen in ihrer Skelettmorphologie gut mit den heutigen Cariamiformes überein, vor allem in der Form des Tarsometatarsus. Die meisten Arten waren kleiner als der Rotfußseriema und eher bodenbewohnend. Im Vergleich zu den Seriemas war die Hinterzehe aber gut entwickelt. Spätere europäische Cariamiformes sind Idiopodia aus dem Miozän und Elaphrocnemus aus dem Eozän und Oligozän. Elaphrocnemus unterscheidet sich aber in einigen Aspekten von Dynamopterus, die ihn außerhalb der Idiornithidae, Terrorvögel und Seriemas stellen. Der Tarsometatarsus von Elaphrocnemus ist kürzer als der von Dynamopterus und den heutigen Seriemas und der Oberarmknochen weist einen stärkeren delapectoralen Kamm auf, der darauf hindeutet, dass er besser fliegen konnte.
Itaboravis wurde aus dem späten Paläozän Brasiliens beschrieben und weitere Fossilien aus dem mittleren Eozän bis späten Oligozän sind aus Nordamerika bekannt und wurden in die Verwandtschaft der Cariamiformes gestellt. Diese Vögel wurden lange Zeit den Bathornithidae zugeordnet, die in ihrer traditionellen Zusammensetzung mit ziemlicher Sicherheit ein polyphyletisches Taxon sind. Die am besten vertretenen Gattungen dieser Familie sind Bathornis („Neocathartes“) aus dem mittleren Eozän von Wyoming und Paracrax aus dem frühen Oligozän von South Dakota. Bathornis scheint flugunfähig gewesen zu sein, wie die Form der Flügelknochen und die langen Beine belegen. Im Unterschied zu den Terrorvögeln und Seriemas ist die Hinterzehe von Bathornis gut ausgebildet.
Die Paracrax-Arten waren große bis sehr große Vögel, und die größte Art, Paracrax gigantea, wurde mehr als doppelt so groß wie die heutige Rotfußseriema. Die Skelettmorphologie, wie die Flügelknochen und das Brustbein, von Paracrax unterscheidet sich stark von der von Bathornis. Das reduzierte Brustbeinkiel von Paracrax ähnelt dem von Hoatzinen, was auf die Existenz eines großen Kropfes und eine vegetarische Ernährung hinweisen könnte. Obwohl vorgeschlagen wurde, dass Paracrax eine Klade mit den Idiornithidae und Seriemas bildet, zeigt sein Oberarmknochen eine größere Ähnlichkeit mit dem von Elaphrocnemus und Hoatzins.
Offensichtlich haben die Cariamiformes ihre Flugfähigkeit mehrmals unabhängig voneinander verloren, und zwar bei den südamerikanischen Terrorvögeln, einigen europäischen Taxa und dem nordamerikanischen Bathornis. Eine Voraussetzung für den Verlust der Flugfähigkeit bei diesen Vögeln ist der Umstand, dass sie auf das Laufen spezialisiert waren und am Boden nach Nahrung suchten, was die Aufgabe einer fliegenden Lebensweise in Umgebungen mit geringerem Feinddruck begünstigte. Höchstwahrscheinlich war die Stammart der Gruppe, zu der Bathornis, Dynamopterus, die Terrorvögel und die heute lebenden Seriemas gehören, bereits ein langbeiniger Landvogel mit schwachen Flugfähigkeiten. Ein mögliches Kandidatentaxon für einen solchen frühen Stammgruppenvertreter ist Gradiornis aus dem späten Paläozän Deutschlands, das allerdings nur von wenigen Knochen bekannt ist.
Die Ordnung der Cariamiformes belegt, wie reichhaltig einige Gruppen in ihrer Evolutionsgeschichte waren und gleichzeitig wie fragmentarisch der Fossilbeleg bei vielen anderen Vogelgruppen ist. Umso wichtiger sind molekularbiologische Vergleiche zur Ermittlung der Verwandtschaftsbeziehungen innerhalb der Vögel.
Falconiformes

Trotz ihrer ähnlichen Erscheinung gehören Falken nicht zu den Greifvögeln, sondern bilden eine eigene Ordnung, die enger mit den Papageien und Sperlingsvögeln verwandt sind. Früher wurden sie tatsächlich mit den Habichtartigen in die Ordnung Falconiformes gestellt, bis molekularbiologische Untersuchungen belegten, dass ihre oberflächliche Ähnlichkeit Resultat einer konvergenten Evolution ist: Wenn nicht näher miteinander verwandte Gruppen ähnliche ökologische Nischen besetzen führt die natürliche Selektion zu oberflächlich ähnlichen Merkmalsausprägungen. Gleichzeitig zeigt uns die Evolutionsbiologie einiges: Divergierende Evolutionslinien können vom gemeinsamen Vorfahren so stark abweichen, dass nach zig Generationen evolutiven Wandels ein gemeinsamer Ursprung kaum zu erkennen ist (Winkler et al. 2015, 2020b, Del Hoyo 2020, Del Hoyo et al. 1994).
Welche Merkmale zeichnen die Falken als Familie aus? Grundsätzlich sind Falken leichtgewichtig und ihre Flugsilhouette weist lange sichelförmige Flügel aus, meist ohne eine Fingerung der Flugfedern, wie sie für Greifvögel typisch ist. Die Brustmuskulatur mach zwischen 12 und 20% des Körpergewichts aus. Falken wechseln ihr Gefieder in einer jährlichen Vollmauser, bei der das Klein- und Großgefieder vollständig ersetzt wird. Die Federn werden dabei in einer bestimmten, für die Familie typischen, Reihenfolge ersetzt: Sie beginnt mit der vierten Handschwinge im Flügel und setzt sich von da an zu beiden Seiten nacheinander fort. Das Gefieder weist oft braune, graue oder weiße Farben auf und ist oft gebändert oder fleckig.
Sie haben für fleischfressende Vögel den typisch gebogenen Schnabel. Anders als Greifvögel, die ihre Beute mit den Fängen töten, benutzen Falken dazu den Schnabel. Kleine Schnäbel und starke Kiefermuskeln ermöglichen deshalb einen kraftvollen Biss (Sustaita 2008). Am Oberschnabel befindet sich eine typische Ausformung, der Falkenzahn genannt wird, und den Vogel beim Biss in den Nacken des Beutetieres unterstützt. Falken sind unglaublich schnelle Flieger: Wanderfalken (Falco peregrinus) wurden beim Sturzflug mit Geschwindigkeiten von 320 km/h gemessen, was sie zu den schnellsten Lebewesen auf der Erde macht; der schnellste gemessene Sturzflug erreichte eine Geschwindigkeit von 390 km/h. Falken bauen keine Nester. Die Brut findet in Nestern anderer Vogelarten oder in einfachen Mulden an Felswänden, Gebäuden oder auf Bäumen statt. Auch das unterscheidet sie von den Greifvögeln. Falken sind tagaktive Jäger. Im Gegensatz zu Adlern oder Bussarden ist ihre Anatomie auf den aktiven Flug hin ausgerichtet und nicht optimal zum Nutzen von Aufwinden geeignet. Dies führt dazu, dass die meisten Falkenarten ihre Beute im aktiven Flug suchen. Wird diese entdeckt, wird sie auch über weite Strecken hin angeflogen und verfolgt. Typisch für einige Falken ist der Rüttelflug, welcher besonders beim Turmfalken (Falco tinnunculus) zu beobachten ist. Beim Rüttelflug bleibt die Position des Vogels in Bezug auf den Boden unverändert. Der Körper wird dabei aufrecht gehalten, die Flügel stehen in einem so großen Anstellwinkel, dass der Flügelschlag keinen Vortrieb, sondern stattdessen hohen Auftrieb erzeugt. Auftrieb und Schwerkraft befinden sich im Gleichgewicht, das Tier verliert daher im Rüttelflug nicht an Höhe. Außer bei Falken ist diese Art des Fliegens auch beim Mäusebussard oder bei Möwen und Eisvögeln zu beobachten. Der Schwirrflug der Kolibris unterscheidet sich durch andere Flügelbewegungen vom Rüttelflug. Falken haben 15 Halswirbel. Ein Turmfalke beispielsweise, der von einer Warte aus nach Beute ausspäht, kann seine Halswirbelsäule um 180° drehen. Schon aufgrund seiner Augenstellung beträgt sein Blickfeld 220°, ohne dass er den Hals auch nur drehen muss.
Die kleinsten Falken, das Finkenfälkchen (Microhierax fringillarius) und das Weißscheitelfälkchen (Microhierax latifrons), beide aus Südostasien, wiegen nur 35 Gramm und erreichen Flügellängen von etwa 10 cm. Der größte, der im hohen Norden lebende Gerfalke (Falco rusticolus), erreicht ein Gewicht von bis zu 1,7 kg und Flügelspannweiten zwischen einem und 1,30 Metern.
Insgesamt gibt es 65 Falkenarten, die – je nach Autor – in 10 bis 12 Gattungen eingeteilt werden. Nach neuesten Studien werden zwei Unterfamilien anerkannt. Falken kommen fast weltweit vor, fehlen aber z. B. in vielen ozeanischen Inseln und im Regenwald des Kongobeckens. Die größte Artenvielfalt findet sich in Südamerika und den Savannen Afrikas.
Die Unterfamilie Herpetotherinae besteht aus zwei Gattungen: die Lachfalken (Herpetotheres) mit einer Art (Herpetotheres cachinnans) und den Waldfalken (Micrastur) mit sieben Arten. Sie alle kommen in den Regenwäldern von Mittel- und Südamerika vor.
Die Unterfamilie Falconinae hat zwei Gattungsgruppen: die Polyborini und die Falconini. Die Polyborini sind wie die Herpetotherinae nur auf Südamerika beschränkt. Sechs Gattungen mit 11 Arten sind bekannt, 5 Gattungen mit 10 Arten gehören zu den sogenannten Geierfalken oder Karakaras, von deinen eine Art, der Schopfkarakara (Caracara plancus), bis in den Süden der USA vorkommt. Geierfalken sind im Gegensatz zu ihren Verwandten keine schnellen Flieger und in Bezug zu ihrer Nahrung auch Opportunisten, die auch, wie der Name vermuten lässt, Aas fressen.
Neben den fünf eigentlichen Geierfalken-Gattungen gehört auch der Tropfen-Zwergfalke als einziger Vertreter seiner Gattung (Spiziapteryx circumcinctus) zu den Polyborini. Seinem Äußeren nach ähnelt er den Falken, ist aber mit den Geierfalken näher verwandt. Die Geierfalken wurden traditionell als eigene Unterfamilie innerhalb der Falken geführt. Die nähere Verwandtschaft zum Tropfen-Zwergfalken und dann Lach- und Waldfalke die basalste Falkenarten sind, hat zur Aufgabe der eigenständigen Unterfamilie der Geierfalken geführt (Griffith et al. 2004, Fuchs et al. 2012).
Die Gattungsgruppe der Falconini umfasst vier Gattungen. Die Gattung Polihierax hat eine Art, den in Afrika vorkommenden Halsband-Zwergfalken (Polihierax semitorquatus). Die Schwestergruppe dieser Gattung sind die in Südostasien vorkommenden Eigentlichen Zwergfalken (Microhierax) mit 5 Arten; sie sind die kleinsten Vertreter aller Falken. Der Langschwanz-Zwergfalke (Neohierax insignis) wird in eine eigene Gattung, Neohierax, gestellt und kommt in Südostasien vor (Fuchs et al. 2014).
Die Gattung Falco hat 39 Arten, wozu auch die in Mitteleuropa brütenden Arten Turmfalke (Falco tinnunculus), Baumfalke (Falco subbuteo), Rotfußfalke (Falco vespertinus), Würgfalke (Falco cherrug) und Wanderfalke (Falco peregrinus) gehören. Der Merlin (Falco columbarius) brütet in der nördlichen Holarktis und nordeuropäische Populationen sind im Herbst Durchzügler in Mitteleuropa.
Der Turmfalke (Falco tinnunculus) wird in mehrere Unterarten aufgeteilt, die in Europa, Asien und Afrika vorkommen. Aufgrund zahlreicher Ringfunde weiß man, dass Turmfalken sowohl Stand-, Strich- als auch ausgeprägte Zugvögel sein können. Ihr Zugverhalten ist im Wesentlichen von dem Nahrungsangebot geprägt, das ihnen in ihren jeweiligen Brutarealen zur Verfügung steht. Die Brutvögel Deutschlands, der Niederlande und Belgiens sind überwiegend Stand- und Strichvögel.
Wanderfalken (Falco peregrinus) haben eine fast weltweite Verbreitung und fehlen in den tropischen Wäldern Südamerikas und Afrikas und in den Steppenregionen Asiens. Sie meiden außerdem die Wüsten Amerikas, Afrikas und Australiens. Wanderfalken zeigen eine deutliche Größenzunahme nach Norden. Außerdem sind die Wanderfalken in feuchten Klimazonen dunkler als in trockenen. Es gibt mehrere Unterarten des Wanderfalken, deren genaue Anzahl aber unklar ist, da es zwischen den bis zu 19 Unterarten fließende Übergänge gibt.
Eine relativ hohe Artenvielfalt der Gattung Falco findet sich in den afrikanischen Savannen, insgesamt kommen dort über ein Dutzend Arten vor, sowie weitere Arten auf Madagaskar und den Inseln des indischen Ozeans. Bekannte afrikanische Arten sind Lanner-, Grau-, Felsen-, und afrikanischer Baumfalke (F. biarmicus, F. ardosiaceus, F. rupicolus, F. cuvierii). Der Taitafalke (F. fasciinucha) kommt nur in einem begrenzten Gebiet in Kenia vor.
Fossile Falken
Es gibt eine Reihe fossiler Falken, von denen zu den frühesten Stammgruppen Masillaraptor aus dem Eozän der Grube Messel gehört. Masillaraptor hat einen langen Schnabel mit einem geraden Dorsalkamm, der kurz vor der Spitze gebogen ist. Die Beine sind lang und die Krallen eher schwach. Wie bei den heutigen Falken und Habichtartigen sind einige Zehenglieder verkürzt (Mayr 2006c, 2009b). Mutmaßliche Falken wurden aus dem Eozän von England und der Seymour-Insel (Antarktis) beschrieben. Diese Belege bestehen aber nur aus Tarsometatarsi und ihre genaue Einordnung ist nicht in jedem Fall sicher (Mayr 2006, 2009, 2016, Mayr & Kitchener 2022, Noriega et al. 2011, Becker 1987, Fuchs et al. 2015, Olson 2008).
Basierend auf den phylogenetischen Beziehungen der modernen Falken dürften diese ihre Ursprünge in Südamerika haben, was auch durch Fossilfunde belegt ist. So findet man aus dem Miozän Argentiniens die Gattung Thegornis, die Ähnlichkeiten mit den basalen Lachfalken hat (Noriega et al. 2011). Fossile Geierfalken treten erstmal im späten Miozän Argentiniens auf (Cenizo et al. 2012). Der früheste Vertreter der Falconini ist Pediohierax aus dem mittleren Miozän von Nebraska (Becker 1987b). In der Alten Welt traten die Falconini erstmals im späten Miozän in Europa und Asien auf, und alle diese Arten wurden der heutigen Gattung Falco zugeordnet (Boev 2011; Li et al. 2014b).
Die Fossilfunde von Falken deuten also darauf hin, dass die Kronengruppe dieser Familie im Oligozän Südamerikas entstand, von wo aus sich die Falken im mittleren bis späten Känozoikum über Nordamerika nach Europa ausbreiteten. Falken der Klade Herpetotherinae sind stärker an ein Leben in bewaldeten Regionen angepasst als die Arten der Polyborini und Falconini, und die Ausbreitung der Falken nach Norden könnte auf die Ausbreitung offener Landschaften in der nördlichen Hemisphäre im mittleren Känozoikum zurückzuführen sein (Fuchs et al. 2015). Die Entstehung der modernen Falken im Oligozän ist insofern spannend, weil der gemeinsame Vorfahre der Kronengruppe der Psittacopasseres (also die Papageien plus Sperlingsvögel), welche die Schwestergruppe der Falconiformes sind, viel älter sind. Wenn die Falken korrekt als Schwestergruppe der Papageien und Sperlingsvögel eingeordnet werden, müssen sie im frühen Eozän oder Paläozän vor 50 – 60 Mio. Jahren entstanden sein. Doch nur eine Linie aus dem Oligozän (vor etwa 30 Millionen Jahren) ist der Vorfahre aller heutigen Falken, während alle anderen Falken-Linien aus den vorangegangenen 20-30 Millionen Jahren ausgestorben sind. Leider ist die Fossilüberlieferung der Falken zu spärlich, um ihre frühe Evolutionsgeschichte zu rekonstruieren.
Falken sind also mit den Papageien näher verwandt als mit den Greifvögeln. Dementsprechend werden wir diese Gruppe in der nächsten Episode näher behandeln.
Literatur
Acosta Hospitaleche, C.; Jones, W. (2024). Insights on the oldest terror bird (Aves, Phorusrhacidae) from the Eocene of Argentina. Historical Biology: An International Journal of Paleobiology. 37 (2): 391–399.
Alvarenga, H. (2014). South American and Antarctic Continental Cenozoic Birds — Paleobiogeographic Affinities and Disparities. Ameghiniana. 51 (3). Asociacion Paleontologica Argentina: 266.
Alvarenga, H., Chiappe, L., Bertelli, S. (2011): Phorusracids: The Terror birds. In: Gareth Dyke, Gary Kaiser (Hrsg.): Living Dinosaurs: The History and evolution of modern birds. John Wilex & Sons, 2011, ISBN 978-0-470-65666-2, S. 187–203
Alvarenga, H.; Höfling, E. (2003). Systematic revision of the Phorusrhacidae (Aves: Ralliformes). Papéis Avulsos de Zoologia. 43 (4): 55–91.
Angst, D.; Buffetaut, E.; Lécuyer, C.; Amiot, R. (2013). ‚Terror Birds‘ (Phorusrhacidae) from the Eocene of Europe Imply Trans-Tethys Dispersal. PLOS ONE. 8 (11) e80357.
Becker, J. J. (1987). Revision of „Falco“ ramenta Wetmore and the Neogene evolution of the Falconidae. Auk 104(2):270-276
Cenizo, M. M. (2012). Review Of The Putative Phorusrhacidae From The Cretaceous And Paleogene Of Antarctica: New Records Of Ratites And Pelagornithid Birds. Polish Polar Research. 33 (3): 239–258.
Cracraft, J. (1968). A review of the Bathornithidae (Aves, Gruiformes), with remarks on the relationships of the suborder Cariamae. American Museum Novitates (2326): 1–46.
Cracraft, J., et al. (2004): Phylogenetic relationships among modern birds (Neornithes): towards an avian tree of life. In J. Cracraft and M. Donoghue (eds.): Assembling the Tree of Life, 468– 489. Oxford: Oxford University Press.
Degrange, F. J. (2020). A revision of skull morphology in Phorusrhacidae (Aves, Cariamiformes). Journal of Vertebrate Paleontology. 40 (6) e1848855.
Degrange, F. J.; Tambussi, C. P.; Taglioretti, M. L.; Dondas, A.; Scaglia, F. (2015). A new Mesembriornithinae (Aves, Phorusrhacidae) provides new insights into the phylogeny and sensory capabilities of terror birds. Journal of Vertebrate Paleontology. 35 (2) e912656.
Degrange, F. J., Tambussi, C. P., Witmer, L. M. (2025): Reversing the Trend: The Evolution of Cranial Akinesis in the Terror Birds (Cariamiformes, Phorusrhacidae). Fossil Studies 3, S. 12,
Del Hoyo, J. (2020): All the Birds of the World. Lynx Nature Books
Del Hoyo, J.; Elliot, A.; Sargatal, J., eds. (1994): Handbook of the Birds of the World, volume 2, New World Vultures to Guineafowl, Lynx Edicions, Barcelona.
Del Hoyo, J.; Elliot, A.; Sargatal, J., eds. (1996): Handbook of the Birds of the World, volume 3, Hoatzin to Auks, Lynx Edicions, Barcelona.
Fuchs, J., Johnson, J. A.; Mindell, D. P. (2012). Molecular systematics of the caracaras and allies (Falconidae: Polyborinae) inferred from mitochondrial and nuclear sequence data. Ibis. 154: 520–532.
Fuchs, J., Johnson, J. A.; Mindell, D. P. (2015): Rapid diversification of falcons (Aves: Falconidae) due to expansion of open habitats in the Late Miocene. Molecular Phylogenetics and Evolution Volume 82, Part A:166–182.
Griffiths, C. S., Barrowclough, G. F., Groth, J. G., Mertz, L. (2004): Phylogeny of the Falconidae (Aves): a comparison of the efficacy of morphological, mitochondrial, and nuclear data. Molecular Phylogenetics and Evolution 32: 101–109,
Hackett, S. J. et al. (2008): A phylogenomic study of birds reveals their evolutionary history. Science 320: 1763–1768.
Jarvis, E. D.; et al. (2014): Whole-genome analyses resolve early branches in the tree of life of modern birds. Science. 346 (6215): 1320–1331
LaBarge, T. W.; Gardner, J. D.; Organ, C. L. (2024). The evolution and ecology of gigantism in terror birds (Aves, Phorusrhacidae). Proceedings of the Royal Society B: Biological Sciences. 291 (2021). 20240235. doi:10.1098/rspb.2024.0235
Mourer-Chauviré, C., Tabuce, R., Mahboubi, M., Adaci, M., Bensalah, M. (2011). A phororhacoid bird from the Eocene of Africa. Naturwissenschaften. 98 (10): 815–823.
Mayr, G. (2002). A new specimen of Salmila robusta (Aves: Gruiformes: Salmilidae n. fam.) from the Middle Eocene of Messel. Paleontologische Zeitschrift 76(2/2): 305-316.
Mayr, G. (2005): Old World phorusrhacids (Aves, Phorusrhacidae): a new look at Strigogyps („Aenigmavis“) sapea (Peters 1987). PaleoBios (Berkeley) 25(1): 11-16
Mayr, G. (2006). A new raptorial bird from the Middle Eocene of Messel, Germany. Historical Biology. 18 (2): 99–106.
Mayr, G. (2009). Paleogene Fossil Birds. Heidelberg: Springer
Mayr, G. (2016): Avian Evolution: The Fossil Record of Birds and its Paleobiological Significance. TOPA Topics in Paleobiology.
Mayr, G. (2016a). Osteology and phylogenetic affinities of the middle Eocene North American Bathornis grallator—one of the best represented, albeit least known Paleogene cariamiform birds (seriemas and allies). Journal of Paleontology. 90 (2): 357–374.
Mayr, G. (2017). Cariamiforms and Diurnal Birds of Prey. Avian Evolution: The Fossil Record of Birds and its Paleobiological Significance. Chichester: Wiley-Blackwell. pp. 189–204.
Mayr, G. (2022). Accipitriformes (New World Vultures, Hawks, and Allies), Falconiformes (Falcons), and Cariamiformes (Seriemas and Allies). Paleogene Fossil Birds. Fascinating Life Sciences. Springer Cham. pp. 153–176.
Mayr, G.; Alvarenga, H.; Clarke, J. (2011). An Elaphrocnemus−like landbird and other avian remains from the late Paleocene of Brazil. Acta Palaeontologica Polonica. 56 (4): 679–684
Mayr, G.; Kitchener, A. C. (2022). New fossils from the London Clay show that the Eocene Masillaraptoridae are stem group representatives of falcons (Aves, Falconiformes). Journal of Vertebrate Paleontology. 41 (6) e2083515.
Mayr, G.; Noriega, J.I. (2013). A well-preserved partial skeleton of the poorly known early Miocene seriema Noriegavis santacrucensis (Aves, Cariamidae). Acta Palaeontologica Polonica. 60 (3): 589–598.
Mayr, G.; Richter, G. (2011). Exceptionally preserved plant parenchyma in the digestive tract indicates a herbivorous diet in the Middle Eocene bird Strigogyps sapea (Ameghinornithidae). Paläontologische Zeitschrift. 85 (3): 303–307.
Mayr, G.; Yang, J.; De Bast, E.; Li, C.-S.; Smith, T. (2013). A Strigogyps-like bird from the middle Paleocene of China with an unusual grasping foot. Journal of Vertebrate Paleontology. 33 (4): 895–901
Noriega, J. J.; Areta, J. I.; Vizcaíno, S. F.; Bargo, M. B. (2011). Phylogeny and Taxonomy of the Patagonian Miocene Falcon Thegornis musculosus Ameghino, 1895 (Aves: Falconidae). Journal of Paleontology. 85 (6): 1089–1104.
Olson, S. L. (2008): A New Species of Large, Terrestrial Caracara from Holocene Deposits in Southern Jamaica (Aves: Falconidae). Journal of Raptor Research. 42(4). 265–272
Peters, D. S. (1987): Ein „Phorusrhacidae“ aus dem Mittel-Eozän von Messel (Aves: Gruiformes: Cariamae). Documents des Laboratoires de Géologie de Lyon 99: 71-87.
Prevosti, F. J., Forasiepi, A., Zimicz, N. (2013): The Evolution of the Cenozoic Terrestrial Mammalian Predator Guild in South America: Competition or Replacement? Journal of Mammalian Evolution 20, S. 3–21
Prum, R. O. et al. (2015): A comprehensive phylogeny of birds (Aves) using targeted next-generation DNA sequencing. Nature. 526 (7574): 569–573.
Stidham, T.A; Wang, Y.-Q. (2017). An ameghinornithid-like bird (Aves: Cariamae: Ameghinornithidae?) from the Middle Eocene of Nei Mongol, China. Vertebrata PalAsiatica. 55 (3): 218–226.
Sustaita, D. (2008), Musculoskeletal underpinnings to differences in killing behavior between North American accipiters (Falconiformes: Accipitridae) and falcons (Falconidae). J. Morphol., 269: 283-301. https://doi.org/10.1002/jmor.10577
Tarquini, S. D., Ladevèze, S., Prevosti, F. J. (2022): The multicausal twilight of South American native mammalian predators (Metatheria, Sparassodonta). Scientific Reports 12, S. 1224
Winkler, D. W., Billerman, S. M., Lovette, I. J. (2015): Bird Families of the World: A Guide to the Spectacular Diversity of Birds. Lynx Edicions, Barcelona.
Winkler, D. W., Billerman, S. M., Lovette, I. J. (2020a). Seriemas (Cariamidae), version 1.0. In Birds of the World (S. M. Billerman, B. K. Keeney, P. G. Rodewald, and T. S. Schulenberg, Editors). Cornell Lab of Ornithology, Ithaca, NY, USA. https://doi.org/10.2173/bow.cariam1.01
Winkler, D. W., Billerman, S. M., Lovette, I. J. (2020b). Falcons and Caracaras (Falconidae), version 1.0. In Birds of the World (S. M. Billerman, B. K. Keeney, P. G. Rodewald, and T. S. Schulenberg, Editors). Cornell Lab of Ornithology, Ithaca, NY, USA. https://doi.org/10.2173/bow.falcon1.01
