
Wir befassten uns im letzten Beitrag mit der Zellatmung. Bei der Zellatmung wird aus Zucker mit Hilfe von Sauerstoff Energie in Form von ATP gespeichert. Als Produkte entstehen dabei CO2 und Wasser.
In diesem Beitrag befassen wir uns mit der Photosynthese. Diese ist quasi gesehen die umgekehrte Richtung der Zellatmung: Aus CO2 und Wasser werden Zucker und Sauerstoff.
Die entsprechende Summenformel der Photosynthese lautet: 12 H2O +6 CO2 → C6H12O6 +6 O2 +6 H2O. Eine vereinfachte, verkürzte Formel reduziert die Anzahl der Wassermoleküle, da sie sich mathematisch rauskürzen lassen: 6 H2O +6 CO2 → C6H12O6 +6 O2 (vgl. Abb. 1).

Abb. 1: Bilanz der Photosynthese
Natürlich ist die Photosynthese keine bloße Umkehrung der Zellatmung. Das kann alleine deswegen schon nicht sein, weil Zucker energiereicher ist als CO2 und Wasser. Damit aus diesen beiden Molekülen Zucker gebildet werden kann, muss also zusätzlich Energie verwendet werden. bei der Photosynthese handelt es sich um eine endotherme Reaktion, während die Zellatmung exotherm ist.
Wir erinnern uns: exotherme Reaktionen benötigen wenig Aktivierungsenergie und die Reaktionsprodukte sind am Ende energieärmer als die Ausgangsstoffe, auch als Edukte bezeichnet. Die Aktivierungsenergie wird bei der Zellatmung mittels Enzymen überwunden und die frei gewordene Energie wird in Form von ATP oder Wärme gespeichert.
Bei endothermen Reaktionen braucht es zusätzlich Energie (meist Wärmeenergie) von außen, damit die Ausgangsstoffe miteinander eine Reaktion eingehen. Die entstandenen Reaktionsprodukte sind auf einem höheren Energielevel als die Ausgangsstoffe (Edukte).
Doch woher stammt diese zusätzliche Energie, damit aus CO2 und Wasser Zucker gebildet werden kann? Der Name Photosynthese sagt es bereits: es ist das Licht.
Licht kann aber nicht von Tieren und damit von uns Menschen als Energiequelle zur Produktion von Nährstoffen genutzt werden – auch wenn einige wahnsinnige Esoteriker behaupten Lichtwesen zu sein oder sich von Licht zu ernähren. Wir müssen von außen organisches Material aufnehmen, um uns mit Nährstoffen zu versorgen. Wenn man das tut, spricht von heterotroph.
Organismen, die über die Photosynthese Lichtenergie in chemisch gebundene Energie umwandeln können, sind alle grünen Pflanzen, die Cyanobakterien und einige Bakteriengruppen (phototrophe Bakterien). Letztere betreiben allerdings eine sog. anoxygene Photosynthese, da sie andere Substanzen als H2O als Elektronendonoren verwenden und daher nicht O2 freisetzen. Wir konzentrieren uns auf die oxygene Photosynthese, also jene, die Sauerstoff freisetzen.
Sind Lebewesen in der Lage aus anorganischen Substanzen organische Verbindungen aufzubauen, spricht man von autotroph. Dazu gehören – mit wenigen Ausnahmen – die Pflanzen. Aber warum sind Pflanzen dazu in der Lage?
Hierfür müssen wir uns – bevor wir uns mit der Photosynthese als solches auseinandersetzen – mit den Grundlagen befassen.
Chlorophyll und Chloroplasten
Warum sind Pflanzen eigentlich so grün? Sie verfügen über einen Farbstoff namens Chlorophyll, der ihnen die grüne Farbe verleiht (Chlorophyll heißt auf Deutsch Blattgrün). Pflanzen, Algen und Cyanobakterien besitzen verschiedene Chlorophylltypen. Sie werden als Chlorophyll a, b. c1, c2, d und f bezeichnet. Bei Bakterien, die die anoxygene Photosynthese betreiben, kommen Bakteriochlorophylle vor.
Alle Chlorophylle haben das gleiche Grundgerüst, das aus vier Pyrrolringen, dem sogenannten Porphyringerüst mit mehreren Doppelbindungen und einem im Zentrum komplex gebundenen zweifach positiven Magnesium-Ion Mg2+ besteht (Abb. 2). Porphyrine sind organisch-chemische Farbstoffe mit ringförmigen Kohlenstoffgerüsten. Die in Abb. 2 mit R1-4 abgekürzten Bereiche sind die Reste der verschiedenen Chlorophylle. Außerdem haben die Chlorophylle außer c1 und c2 weisen zusätzlich einen Phytolrest auf (Abb. 2. 2 links im Bild).
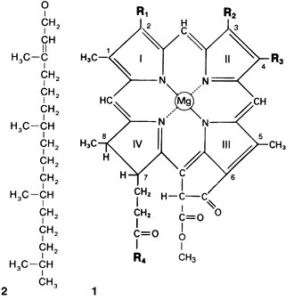
Abb. 2: 1 = Grundgerüst des Chlorophylls; 2= Phytolrest
Die Struktur des Chlorophylls ermöglicht es ihnen das Licht bestimmter Wellenlängen zu absorbieren.
Licht ist ein Teil der elektromagnetischen Wellen. Beispiele für elektromagnetische Wellen sind Radiowellen, Mikrowellen, Wärmestrahlung, Licht, Röntgenstrahlung und Gammastrahlung (Aufzählung nach aufsteigender Frequenz). Im engeren Sinne sind vom gesamten elektromagnetischen Spektrum nur die Anteile als Licht definiert, die für das menschliche Auge sichtbar sind. Im weiteren Sinne werden auch elektromagnetische Wellen kürzerer Wellenlänge (Ultraviolett) und größerer Wellenlänge (Infrarot) dazu gezählt. Weißes Licht ist eine Mischung der Farben aus dem sichtbaren Spektrum das von Violett über Blau, Grün, Gelb und Rot reicht. Diese Farben haben unterschiedliche Wellenlängen zwischen etwa 400 Nanometern beim Violett und 700 Nanometern beim Rot (Abb. 3).
Abb. 3: elektromagnetische Wellen.
Wenn Licht auf einen Gegenstand oder eine Substanz fällt und die Lichtstrahlen nicht reflektiert werden, dann werden sie absorbiert. Sie werden sozusagen „verschluckt“.
Seine Struktur erlaubt es, dass Chlorophyll Licht der grünen Wellenlänge reflektiert (daher erscheint es uns als grün), Licht der roten und blauen Wellenlänge absorbiert (also verschluckt).
Die verschiedenen Chlorophyllmoleküle haben Licht abgewandelte Absorptionsspektren, absorbieren Licht also etwas unterschiedlich. Abb. 4 zeigt die Absorptionsspektren von Chlorophyll a und b.
Zusammen absorbieren Chlorophyll a und b hauptsächlich im blauen Spektralbereich (400–500 nm) sowie im roten Spektralbereich (600–700 nm). Im grünen Bereich hingegen findet keine Absorption statt, so dass grünes Licht gestreut wird, was Blätter grün erscheinen lässt.
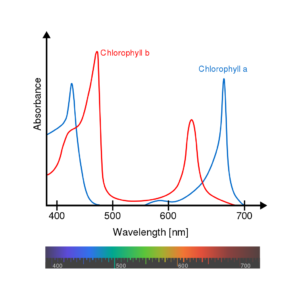
Abb. 4: Absorptionsspektrum von Chlorophyll a und b.
Chlorophylle spielen bei der Photosynthese eine wichtige Rolle, denn sie sind in der Lage die absorbierte Energie des Lichts weiterzuleiten, also quasi als photochemischer Katalysator, welches Elektronentransportketten initiiert.
Damit solche Elektronentransportketten, wir haben ja schon welche bei der Zellatmung kennengelernt, effizient funktionieren, muss das Chlorophyll in Lichtsammelkomplexen organisiert sein. Chlorophyll „schwimmt“ nicht einfach so frei in der Pflanzenzelle herum, sondern befindet sich konzentriert in einem für sie typischen Organell: den Chloroplasten (Abb. 5).
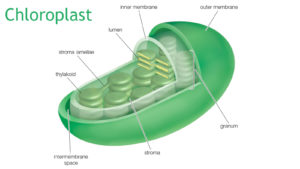
Abb. 5: Chloroplast
Der strukturelle Aufbau der Chloroplasten gleicht dem der Cyanobakterien. Chloroplasten besitzen (fast immer) eine eigene DNA (Chloroplasten-DNA, abgekürzt cpDNA oder ctDNA) zusammen mit eigenen Ribosomen. Chloroplasten haben eine Doppelmembran. Innerhalb er inneren Membran befindet sich die Chloroplastenmatrix, auch Stroma genannt. Eingebettet in diese Matrix liegt ein internes Membransystem (die Thylakoidmembran). Dieses Thylakoid-Membransystem kann geldrollenartig übereinandergestapelt sein, was man als Granum bezeichnet. In den Membranen der Thylakoide sind verschiedene Pigmente eingelagert, vor allem der grüne Farbstoff Chlorophyll. Besonders viel davon findet sich in den Membranen der Grana, weshalb diese intensiv grün gefärbt erscheinen. Hier findet die Photosynthese statt.
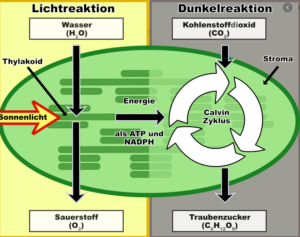
Die Photosynthese kann in die beiden Teilprozesse Lichtreaktion und Dunkelreaktion untergliedert werden. Die Lichtreaktion findet in der Thylakoidmembran, die Dunkelreaktion im Stroma statt. Im Rahmen der Lichtreaktion wird die Energie bereitgestellt.
In der Dunkelreaktion wird diese Energie dazu eingesetzt, um aus CO2 energiereiche organische Verbindungen zu synthetisieren.
Wir werden uns beide Reaktionen: Lichtreaktion und Dunkelreaktion näher betrachten.
Lichtreaktion
Die Lichtreaktion läuft in der Thylakoidmembran der Chloroplasten ab. Diese enthält zwei große Proteinkomplexe die man als Photosysteme 1 und 2 bezeichnet (Abb. 6). Sie sind mit der Fähigkeit ausgestattet, Sonnenergie einzufangen und umzuwandeln. Weiterhin befinden sich zwei Proteinkomplexe, die denen der Atmungskette der Mitochondrien ähneln: ein Cytochrom-b6-f-Komplex und die Chlorplasten-ATP-Synthase.
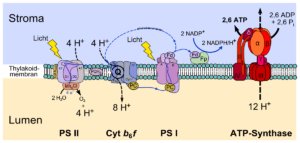
Abb. 6: Feinbau der Thylakoidmembran
Ein Photosystem setzt sich aus einem sogenannten Antennenkomplex und aus einem Reaktionszentrum zusammen. Der Antennenkomplex besteht aus ca. 30 Proteinen, die mit Pigmentmolekülen verbunden sind. Sie werden durch das Licht in einen energiereichen, angeregten Zustand angehoben. Eine Photosynthese durch einfache Pigmente wäre relativ ineffizient, da diese dem Licht nur eine geringe Fläche entgegenstellen würden und zudem nur in einem engen Wellenlängenbereich absorbieren würden. Durch die Anordnung von chlorophyllhaltigen Lichtsammelkomplexen zu Antennen um ein gemeinsames Reaktionszentrum wird sowohl der Querschnitt vergrößert, als auch das Absorptionsspektrum verbreitert. Die eng benachbarten Chromophore in den Antennen geben die Lichtenergie von einem Pigment zum anderen weiter. Durch Exzitonentransfer kann diese Energie an das Reaktionszentrum weitergeleitet werden. Das Reaktionszentrum der Photosysteme enthält zwei Chlorophylle, die als primärer Elektronendonator fungieren. Durch die Lichtenergie wird eine Elektronentransportkette in Gang gesetzt.
Beide Photosysteme unterscheiden sich in ihren Details und Absorptionsspektren.
Das Photosystem I enthält insgesamt ca. 200 Moleküle Chlorophyll a und b sowie 50 Carotine. Das Reaktionszentrum des Photosystems I hat ein Absorptionsmaximum bei einer Wellenlänge von 700 nm, es wird deshalb auch als „P700“ bezeichnet (Abb. 7).
Das Photosystem II enthält insgesamt ca. 250 Moleküle Chlorophyll a und b sowie ca. 110 Carotinoide. Das Reaktionszentrum des Photosystems II hat ein Absorptionsmaximum bei 680 nm („P680“, Abb. 8).
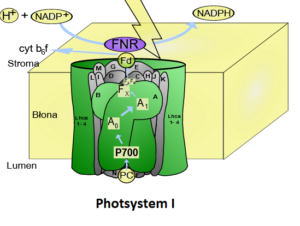
Abb. 7: Photosystem I
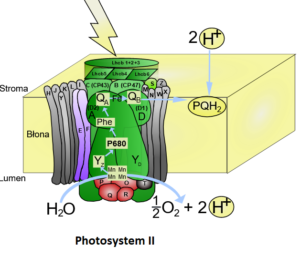
Abb. 8: Photsystem II
Das Chlorophyllmolekül besteht aus Pyrrolringen mit konjugierten Doppelbindungen und einem zentralen Magnesiumion. Konjugierte Doppelbindungen bedeuten, dass sich im Kohlenstoffgerüst Einfachbindungen und Doppelbindungen abwechseln. Bei solchen sich abwechselnden Einfach- und Doppelbindungen bei Ringstrukturen finden sich delokalisierte Elektronen. D. h. ein oder mehrere Elektronen eines Moleküls sind nicht genau lokalisierbar, sondern über die einzelnen Atome verteilt.
Vereinfacht ausgedrückt: Sie können sich nicht entscheiden an welcher Stelle der Bindung sie bleiben sollen und wandern hin und her, sodass sich in einem Kohlenstoffring mit konjugierten, also abwechselnden, Doppelbindungen die Position der Einfach und Doppelbindungen ändert. Man spricht auch von mesomeren Grenzstrukturen. Dies ist am Beispiel des Benzols in Abb. 9 dargestellt.
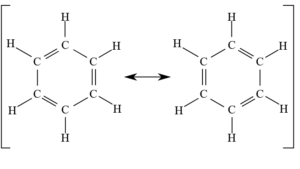
Abb. 9: mesomere Grenzstrukturen beim Benzol.
Solche mesomeren Grenzstrukturen sind beim Chlorophyll ebenso vorhanden und das hat entsprechende Konsequenzen. Absorbiert ein Chlorophyllmolekül ein Lichtquant, führt die Energie des Lichtquants dazu, dass eines der delokalisierten Elektronen angeregt wird und von einer energieärmeren Schale bzw. Orbital (wir erinnern uns an das Schalenmodell der Atome) in ein energetisch höheres angehoben wird. Dieses angeregte Elektron im Chlorophyllmolekül strebt aber danach auf seinen Grundzustand zurückzukehren. Welche Möglichkeiten hat nun dieses angeregte „ADHS-Elektron“ seinen Grundzustand zu erreichen?
Folgende drei Möglichkeiten gibt es:
- Umwandlung der zusätzlichen Energie in Wärme
- Direkte Übertragung der Energie – aber nicht des Elektrons – auf ein anderes Chlorophyllmolekül in der Nachbarschaft
- Übertragung des angeregten Elektrons auf einen Elektronenakzeptor, dann Rückkehr des nun positiv geladenen Chlorophyllmoleküls in seinen Grundzustand durch Aufnahme eines anderen, nicht angeregten Elektrons.
Die letzten beiden Fälle treten auf, wenn Chlorophyll an Proteinkomplexe gebunden ist. Das trifft auf die Photosysteme zu. Photosysteme II und I sind in Serie geschaltet und durch eine Elektronentransportkette verbunden. Werden die Redoxpotentiale aller an der Reaktion beteiligten Redoxpartner aufgetragen, ergibt sich eine Art Zick-Zack-Verlauf, der an ein gedrehtes „Z“ erinnert (Abb. 10).
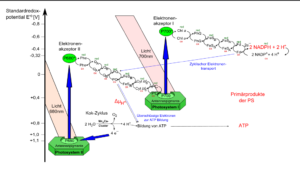
Abb. 10: Ablauf der Lichtreaktion
Beginnen wir beim Photosystem II (diesen Namen hat er übrigens, weil er als zweites entdeckt wurde!). Die Farbstoffpigmente der Lichtsammelkomplexe transportieren die Lichtenergie weiter, bis sie das Reaktionszentrum erreicht haben.
In diesem Reaktionszentrum absorbiert das zentrale Chlorophyll-Paar schließlich so viel Energie, dass es das angeregte Elektron tatsächlich auf einen Elektronenakzeptor abgibt.
Das positiv geladene Chlorophyll-Molekül ist nun ein sehr starker Elektronenakzeptor (Oxidationsmittel), da er wieder zur Ladungsneutralität gelangen möchte. Deshalb entzieht er einem Wassermolekül das fehlende Elektron. Wasser wird aus diesem Grund durch einen wasserspaltenden Proteinkomplex in Sauerstoff, zwei Elektronen und zwei Wasserstoffprotonen (H+) gespalten.
Hier fällt eines auf: der produzierte Sauerstoff stammt nicht aus dem CO2 – der bei der Lichtreaktion noch keine Rolle spielt, sondern aus dem H2O, also dem Wasser!
Das abgegebene Elektron wird nun über eine Kette an Redoxsystemen (primärer Elektronenakzeptor => Plastochinon => Cytochrom-b6–f-Komplex => Plastocyanin) an das Photosystem I weitergeleitet. Jedes Redoxsystem besteht aus einem Elektronenakzeptor, der ein Elektron aufnimmt und dadurch zum Elektronendonator wird. Der Elektronendonator gibt dieses Elektron dann an ein weiteres Redoxsystem mit geringerem Energiegehalt ab. Bei diesem Elektronentransport wird Energie frei, wodurch die Redoxsysteme Protonen in den Thylakoidinnenraum „pumpen“. Dazu kommen wir aber später noch einmal genauer.
Weiter geht es am Photosystem I: Das Elektron, das vom Photosystem II über die Elektronentransportkette weitergeleitet wurde, erreicht nun das Photosystem I. Es erfolgt auch hier analog zum Photosystem II eine Anregung des sichtbaren Lichts und eine Weitergabe der Energie bis zum Reaktionszentrum.
Dort besitzt das spezielle Chlorophyll a Molekül ein Absorptionsmaximum von 700 nm. Das spezielle Chlorophyll Molekül gibt sein Elektron nun genauso wie beim PSII durch Lichtanregung ab und überträgt es auf ein weiteres Redoxsystem. Dadurch entsteht wieder eine Elektronenlücke am Chlorophyll-Molekül, die über das in der Transportkette vermittelte Elektron geschlossen werden kann.
Das letzte „Glied“ der Elektronentransportkette ist ein Enzym – die NADP+– Reduktase. Das kann NADP+ nun durch Aufnahme von 2 Elektronen und einem Wasserstoffproton (H+) zu NADPH reduzieren. NADP+ steht für Nicotin-Amid-Adenin-Dinucleotid-Phosphat, ein Coenzym, das mit dem NAD+ der Zellatmung verwandt ist.
Bildung von ATP
Wie wir bereits aus der Atmungskette der Zellatmung wissen, veranlasst die Elektronentransportkette die Bildung des Energieträgers ATP. Dafür ist ein Proteinkomplex zuständig – die ATP-Synthase. Dieser ist am Ende der Elektronentransportkette hinter dem Photosystem I in der Thylakoidmembran lokalisiert.
Beim Elektronentransport wird Energie frei, die die beteiligten Redoxsysteme zum Transport von Wasserstoffionen (H+) aus dem Stroma in den Thylakoidinnenraum nutzen. Das hat zur Folge, dass einerseits die H+-Konzentration im Stroma abnimmt und dadurch der pH-Wert ansteigt. Andererseits kommt es zu einer hohen H+-Ansammlung im Innenraum, wodurch auch der pH-Wert sinkt. Dadurch entsteht ein Konzentrationsunterschied (chemisches Potential) – in diesem Fall ein Protonengradient, da wir es hier mit Protonen zu tun haben. Zusätzlich entsteht ein Spannungsunterschied (elektrisches Potential), da die Stromaseite nun negativ und der Innenraum positiv geladen sind.
Biomembranen wie die Thylakoidmembran bestehen aber aus einer Doppelschicht aus Phospholipiden und bilden dadurch eine Barriere für die geladenen Wasserstoffprotonen. Sie sind also im Thylakoidinnenraum „gefangen“. Sie können nur durch ein Kanalprotein – die ATP-Synthase – zurück ins Stroma diffundieren, um den Konzentrations- und Ladungsunterschied auszugleichen. Daran ist die Synthese von ATP – der universellen „Energiewährung“ unserer Zellen – gekoppelt.
Der Protonenrückstrom erzeugt Energie. Das kannst du dir wie bei einem Wasserkraftwerk vorstellen, bei dem Wasser hinter einer Staumauer angestaut wird. Dieser Rückstau wird verwendet, um die Turbinen zur Rotation zu bringen, um Strom zu generieren. Diese Turbine ist unserem Fall die ATP-Synthase. Sie koppelt die Diffusion der Protonen, wie ihr Name vermuten lässt, mit der Synthese von ATP aus ADP und einer Phosphatgruppe. Da in unserem Fall Lichtenergie zu dieser Phosphorylierung führt, kannst du den Prozess auch als Photophosphorylierung bezeichnen.
Zyklischer Elektronentransport
Neben dem gerade kennengelernten linearen Elektronentransport, auch als nichtzyklischer Elektronentransport bezeichnet, läuft unter bestimmten Bedingungen auch ein zyklischer Elektronentransport ab. Dieser besitzt, wie der Name vorgibt, einen kreisförmigen Reaktionsweg. Wichtig ist, dass du dir merkst, dass dieser Elektronentransport ausschließlich zur ATP-Bildung dient. Es entsteht weder Sauerstoff noch NADPH. Er dient also nur zur Energiegewinnung.
Einer der Gründe für diesen Transportweg, der auf den ersten Blick eher ineffizient erscheint, ist, dass oft mehr ATP benötigt wird, als über den nicht zyklischen Reaktionsweg bereitgestellt werden kann. Der zyklische Elektronentransport beginnt und endet im Photosystem I. Das durch Lichtanregung freigesetzte Elektron wird über Redoxsysteme in der Elektronentransportkette zwischen PSII und PSI übertragen und wieder zurück zum Reaktionszentrum des Photosystems I geleitet. Das darin enthaltene Chlorophyll-Molekül kann somit das Elektron aufnehmen (=Reduktion) und kehrt in seine ursprünglich ungeladene Form zurück.
Dabei ist auch ein kleines organisches Molekül beteiligt – das Plastochinon. Es veranlasst den Transport von zwei Wasserstoffprotonen in das Thylakoidlumen. Der daraus entstehende Protonengradient kann dann für die ATP-Synthese verwendet werden.
Die Bruttogleichung der Lichtreaktion kann man folgendermaßen formulieren:
12 H2O + 12 NADP+ + 18 ADP + 18 Pi -> 6 O2 + 12 NADPH + 18 ATP
Calvin Zyklus
Der Calvinzyklus ist ein Teilprozess der Photosynthese, auch Dunkelreaktion genannt. Dunkelreaktion deswegen, weil er auch ohne Licht ablaufen kann. Allerdings ist das etwas irreführend, denn ohne die vorherige Lichtreaktion kann auch keine Dunkelreaktion stattfinden. Die Lichtenergie muss dort nämlich zunächst über einen Umweg in chemische Energie – ATP und NADPH – umgewandelt werden. weiterhin ist der Calvin-Zyklus stark temperaturabhängig. Das kommt daher, dass im Calvin Zyklus viele Enzyme (=Proteine) beteiligt sind, die zu hohen Temperaturen nicht standhalten können (Denaturierung).
Der Calvinzyklus ist – ähnlich dem Zitronensäurezyklus der Zellatmung, eine im Kreis ablaufende Reaktionsfolge, die von verschiedenen Enzymen katalysiert wird. Er wurde nach seinem Entdecker, dem amerikanischen Biochemiker Melvin Calvin bekannt und findet bei Pflanzen im Stroma (=dem Inneren) der Chloroplasten und bei Bakterien im Cytoplasma statt.
Der Calvinzyklus ist die Fixierung des CO2 aus der Luft und dessen Einbau in organische Moleküle.
Der Calvin-Zyklus (Abb. 11) kann in drei Phasen untergliedert werden: Die CO2-Fixierung, die Reduktionsphase und die Regenerationsphase. Das Schema des Calvin Zyklus verläuft folgendermaßen: Kohlenstoffdioxid aus der Luft diffundiert in Pflanzen durch Poren – sogenannte Spaltöffnungen oder Stomata – in die Blätter und von da in die Chloroplasten. Dort findet dann der Calvin Zyklus statt. Verschiedene Enzyme sorgen nun dafür, dass die Kohlenstoffatome des Kohlenstoffdioxid in einer im Kreis ablaufenden Reaktionsfolge verbaut (=Kohlenstoffdioxidfixierung) und daraus stabile energiereiche Verbindungen wie Zuckermoleküle hergestellt werden können.
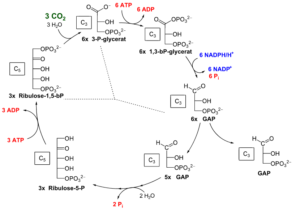
Abb. 12: Calvin-Zyklus
CO2– Fixierungsphase
In der Fixierungsphase verbindet sich ein Kohlenstoffdioxid-Molekül mit einem spezifischen Akzeptormolekül – dem aus 5 Kohlenstoffatomen bestehenden Zucker Ribulose-1,5-bisphosphat (RubP). Diese Reaktion katalysiert ein Enzym mit dem Namen RuBisCO, was für Ribulose-1,5-bisphosphat-Carboxylase-Oxygenase steht.
Hierbei entsteht ein C6– Körper, der instabil ist und sofort in zwei C3–Körper zerfällt. Sie werden auch als 3-Phosphoglycerinsäure (3-PGS) oder 3-Phosphorglycerat bezeichnet. Aus einem Kohlenstoffdioxid-Molekül entstehen also zwei 3-PGS Moleküle.
Reduktionsphase
Im nächsten Schritt – der Reduktionsphase – findet eine chemische Reduktionsreaktion statt, es werden also Elektronen aufgenommen. 3-Phosphoglycerinsäure reagiert zu einer ebenfalls aus 3 Kohlenstoffatomen bestehenden Verbindung namens Glycerinaldehyd-3-phosphat (GAP).
Da es sich um einen energieaufwendigen Prozess handelt, kann dieser nur ablaufen, wenn Energie von außen bereitgestellt wird. Da kommt jetzt die Lichtreaktion ins Spiel. Sie stellt nämlich die aus Lichtenergie gewonnenen Substrate ATP und das NADPH zur Verfügung.
Schauen wir uns nun die Reduktion im Detail an: Sie verläuft in zwei Teilschritten. Zunächst wird die 3-Phosphoglycerinsäure aktiviert, indem ein Enzym eine Phosphatgruppe auf die Säuregruppe überträgt. Die Phosphatgruppe stammt vom Energieträger ATP, der in ADP und Phosphat gespalten wird. Da die Phosphoglycerinsäure nun 2 Phosphatgruppen besitzt, wird sie jetzt als 1,3-Bisphosphoglycerinsäure bezeichnen.
Daraufhin erfolgt die eigentliche Reduktionsreaktion. Die 1,3-Bisphosphoglycerinsäure nimmt nun 2 Elektronen (e–) und ein Wasserstoffproton (H+) von NADPH auf. Dabei wird die zuvor übertragene Phosphatgruppe wieder abgespalten und aus der Carbonsäuregruppe (-COOH) entsteht nun eine Aldehydgruppe (-COH). Wir erhalten eine Verbindung mit dem Namen Glycerinaldehyd-3-phosphat (GAP). Gleichzeitig wird NADPH zu NADP+ oxidiert.
Sowohl NADPH als auch ATP stehen dem Calvin Zyklus durch die vorherige Lichtreaktion zur Verfügung. Deshalb kann die Dunkelreaktion auch ohne Licht nicht lange ablaufen. Gleichzeitig stehen der Lichtreaktion nach ablaufender Dunkelreaktion nun wieder ausreichend ADP und NADP+ zur Verfügung. Diese beiden Reaktionen sind also voneinander abhängig.
Regenerationsphase
Der letzte Reaktionsschritt – die Regenerationsphase – dient dazu, das Akzeptormolekül (Ribulose-1,5-bisphosphat) wiederherzustellen, also zu regenerieren. Dafür werden etwa 5/6 der gebildeten C3–Moleküle Glycerinaldehyd-3-phosphat (G3P) verwendet. Auch hier ist wieder Energie in Form von ATP-Molekülen nötig. Jetzt kann der Zyklus wieder erneut durchlaufen werden, da das Akzeptormolekül wieder zur Verfügung steht.
1/6 des gebildeten Glycerinaldehyd-3-phosphat (G3P) aber verlässt den Zyklus und kann gemeinsam mit einem weiteren Molekül Glycerinaldehyd-3-phosphat (G3P) zu einem aus 6 Kohlenstoffatomen bestehenden Körper reagieren. Nach einigen Umwandlungsreaktionen entstehen daraus die Zucker Glucose (Traubenzucker) oder Fructose (Fruchtzucker). Sie können dann in den Stoffwechsel eingeschleust und dort abgebaut werden. Dadurch gewinnt die Pflanze Energie, die sie zum Beispiel für das Pflanzenwachstum benötigt.
Unter günstigen Bedingungen, also wenn die Pflanzen mehr Einfachzuckermoleküle herstellt, als sie für ihren eigenen Energiebedarf benötigt, können die Zuckermoleküle als Baustoffe für verschiedene Makromoleküle wie Kohlenhydrate, Fette oder Proteine zur Verfügung. Die Kohlenhydrate können im Stroma der Chloroplasten in Stärkekörnern gespeichert und bei Bedarf mobilisiert werden.
Calvin Zyklus Bilanz
Schauen wir uns nun die Bilanz des Calvin Zyklus an: Ein Glucose-Molekül (C6H12O6) besteht aus 6 Kohlenstoffatomen, wofür also 6 Kohlenstoffdioxid-Moleküle benötigt werden. Dafür sind 6 Umdrehungen des Calvin-Zyklus notwendig und ein Energieaufwand von 18 ATP Molekülen und 12 NADPH-Molekülen.
Die Bruttogleichung der Dunkelreaktion kannst du deshalb folgendermaßen formulieren:
Bruttogleichung der Dunkelreaktion
6 CO2+ 12 NADPH + 12 H+ + 18 ATP -> C6H12O6 + 12 NADP+ + 18 ADP + 18 Pi + 6 H2O
