
Im letzten Beitrag befassten wir uns mit der Mitose, bei dem das genetische Material gleichmäßig auf die Tochterzellen weitergegeben wird.
Wir erinnern uns: Menschen haben 46 Chromosomen: 44 Autosomen und die zwei Gonosomen, auch Geschlechtschromosomen genannt (Bei Frauen XX und beim Mann XY). Genauer gesagt haben wir 23 homologe Chromosomenpaare, d. h. jedes Chromosom kommt doppelt vor. Man spricht von diploid. Die Ausnahme stellt natürlich das Y-Chromosom des Mannes dar, welches nicht homolog zum X-Chromosom ist; dennoch bilden beide das Paar der Geschlechtschromosomen. Der diploide Chromosomensatz kommt zustande, weil bei der sexuellen Vermehrung Eizelle und Spermazelle miteinander verschmelzen, ein Vorgang den man als Befruchtung bezeichnet. Eizelle und Spermazelle sind die Keimzellen und diese haben nur einen halben Chromosomensatz, man spricht von haploid. Wie dieser halbe Chromosomensatz entsteht, wird in diesem Beitrag geklärt.
Meiose
Die Meiose lässt sich in zwei Teilabschnitte gliedern: Meiose I und Meiose II.
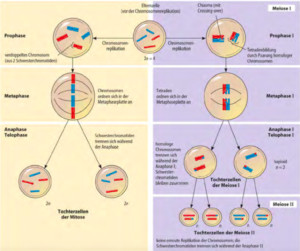
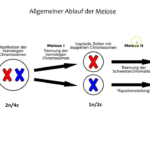
Abb. 1: Unterschied Mitose und Meiose
Der Meiose I geht wie der Mitose eine Interphase mit einer S-Phase voraus, in der jedes Chromosom repliziert wird. Das führt dazu, dass jedes Chromosom aus zwei Schwesterchromatiden besteht, die von Cohesinen zusammengehalten werden.
Die Meiose I beginnt mit einer langen Prophase I, bei der sich die Chromosomen deutlich verändern. Die homologen Chromosomen lagern sich Seite an Seite zu Paaren zusammen, wodurch Tetraden entstehen, also Gruppen aus jeweils vier Chromatiden. Bei der Mitose tritt eine solche Tetradenbildung nicht auf. Die vier Chromatiden stammen von den beiden Partnern eines jeden homologen Chromosomenpaares.
Bei der Mitose verhält sich jedes Chromosom unabhängig von seinem homologen Gegenstück, und seine beiden Schwesterchromatiden werden in der Anaphase zu den beiden entgegengesetzten Polen gebracht. Jeder Tochterzellkern enthält schließlich wieder 2n Chromosomen.
Die Meiose verläuft hier deutlich anders.
In der Meiose I paaren sich in der Synapse die Chromosomen mütterlichen Ursprungs mit ihren väterlichen Gegenstücken. Da jedes Chromosom aufgrund der DNA-Replikation aus zwei identischen Schwesterchromatiden besteht, liegen jetzt Tetraden aus vier sich entsprechenden Chromatiden vor; da die beiden Chromosomen nicht identisch sind, liegen in der Mitte der Tetrade zwei Nicht-Schwesterchromatiden aneinander. Die Paarung mütterlicher und väterlicher homologer Chromosomen tritt bei der Mitose nicht auf.
In der Metaphase I ordnen sich die homologen Chromosomenpaare an der Äquatorialplatte an und werden in der Anaphase I zu den jeweiligen Polen gezogen.
Die Tetraden trennen sich wieder in einzelne Chromosomen, die jeweils aus zwei Schwesterchromatiden bestehen.
Am Ende der Meiose I, genauer der Telophase I, bilden sich zwei Zellkerne; jeder enthält die Hälfte der ursprünglichen Chromosomen (eines von jedem homologen Paar). Da sich die Centromere bis dahin nicht getrennt haben, bestehen diese Chromosomen immer noch aus den beiden Schwesterchromatiden. Die Schwesterchromatiden werden in der Meiose II getrennt, vor der keine DNA-Replikation stattfindet.
Meiose II verläuft im Prinzip wie die Mitose:
In Prophase II kondensieren die Chromosomen, in der Metaphase II ordnen sich die Chromosomen an die Äquatorialplatte und werden in Anaphase II zu den Polen gezogen. Die Schwesterchromatiden trennen sich schließlich und werden eigenständige Tochterchromosomen.
Am Ende der Meiose haben wir also 4 Keimzellen mit dem halben Chromosomensatz. Aber diese vier Zellen sind genetisch nicht identisch.
Um es zusammenzufassen (Abb. 2):
Die Elternzelle ist diploid, das Genom wird repliziert, es entstehen Tetraden (oder anders gesagt: 2n/4c). In der Meiose I ordnen sich die homologen Chromosomenpaare paarweise in der Äquatorialebene an und werden voneinander getrennt. Es entstehen zwei haploide Zellen. Die einzelnen Chromosomen bestehen jedoch noch aus je zwei Chromatiden (1n/2c). In der Meiose II werden die einzelnen Schwesterchromatiden geteilt und es entstehen 4 haploide Zellen. Jedes Chromosom besteht nur noch aus einem Chromatid (1n/1c).
Weil bei der Meiose der Chromosomensatz halbiert wird, spricht man auch von einer Reduktionsteilung.
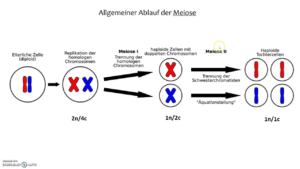
Abb. 2 Meiose
Typen von Keimzellen
Bei einer Meiose entstehen aus einer diploiden Zelle vier haploide. Doch wenn man sich die haploiden Keimzellen, auch Gameten genannt, näher anschaut, sehen sie nicht immer gleich aus. Die beiden sich befruchtenden Gameten gehören zwei verschiedenen, als Geschlechter (Sex) bezeichneten Typen an. Entsprechend gibt es auch zwei unterschiedliche Gametentypen. Die kleineren bzw. beweglichen Gameten sind die männlichen Gameten, die größeren bzw. unbeweglichen Gameten bezeichnet man als weiblich. Die Organismen, die in der Lage sind entsprechende Gameten zu bilden werden als männlich oder weiblich eingestuft. Bei der Befruchtung kann stets nur ein Gamet des einen Typen mit dem anderen verschmelzen. Die Gameten sind also die Ursache der bipolaren Zweigeschlechtlichkeit, was sich durch ihre funktionelle Bedeutung wie auch ihre chemische Struktur, Zelloberfläche etc. ergibt.
Sehen die Keimzellen morphologisch (aber nicht zwingend chemisch oder funktionell!) ähnlich, spricht man von einer Isogamie. Im Falle einer solchen Isogamie werden die beiden, nur an ihren funktionellen Eigenschaften unterscheidbaren Sorten willkürlich als + und – bezeichnet, sofern nicht ein Verhaltensunterschied bei der Einleitung der Befruchtung eine Identifizierung des Geschlechtes erlaubt. Lassen sich die Gameten anhand ihres Aussehens unterscheiden, spricht man von Anisogamie.
Sind beide Gameten begeißelt unterscheidet man zwischen männlichen Mikrogameten und weiblichen Makorgameten.
Ist nur ein Gametentyp begeißelt und damit beweglich, wird dieser – unabhängig von seiner Größe – als männlich definiert.
Sind beide Eigenschaften kombiniert, so sind die kleinen und beweglichen Gameten die männlichen Spermatozoen (bei Tieren Spermien genannt) und die großen unbeweglichen Gameten sind die weiblichen Eier. Diese Kombination bezeichnet man auch als Oogamie. Sie kommt bei allen Tieren (also auch uns Menschen) vor.
Einen Überblick über die verschiedenen Keimzellentypen gibt Abb. 3:
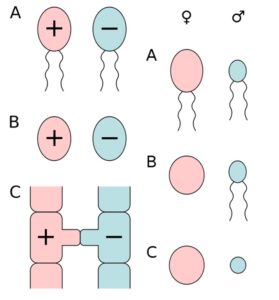
Abb. 3: die unterschiedlichen Gametentypen. Linke Spalte: A) aktiv bewegliche Isogameten, B) unbewegliche Isogamten, C) Gametangiogamie: Befruchtung ohne Gameten bei den Jochpilzen. Rechte Spalte: A) aktiv bewegliche Anisogameten, B) Eizelle und Spermium (Oogamie), C) unbewegliche Anisogameten (Eizelle und Spermatium).
Der Grund weshalb Eizellen übrigens größer sind als Spermien, liegt in einer Sonderfunktion der Meiose bei Eizellen. Hier entstehen zwar auch aus einer diploiden Mutterzelle 4 haploide Zellen. Aber drei dieser haploiden Zellen werden zu sogenannten Polkörpern. Die Teilung der 4 Zellen ist inäqual. Von einer großen Zelle wird nur eine kleine Zelle abgegliedert, die sich dann entweder auflöst oder nochmal teilt. So entsteht eine sehr große Eizelle und 3 sogenannte Polkörper, die für die Befruchtung funktionslos sind (Abb. 4).
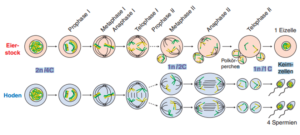
Abb. 4: Spermien- und Eizellenproduktion
Die Produktion unterschiedlicher Gameten geht natürlich einher mit der Ausbildung unterschiedlicher Strukturen ihrer Erzeuger: Das gilt sowohl für primäre Geschlechtsorgane, sowie die sekundären. Im Extremfall bildet sich ein regelrechter Sexualdimorphismus aus, bei dem Männchen und Weibchen so unterschiedlich aussehen, als ob sie anderen Arten angehören könnten (vgl. ALBERTS et al. 2017, CZIHAK 1981, WOLPERT et al. 2007). Da in der Natur also auch immer zwei unterschiedliche Gametentypen pro Art ausgebildet werden, ist auch hier die Einteilung in zwei Geschlechter sinnvoll. Ein drittes Geschlecht würde biologisch nur dann Sinn machen, wenn es triploide Arten gibt, die aus drei unterschiedlichen Keimzellen hervorgehen. Natürlich und das muss gesagt werden: Aus der Bildung der Keimzellen lassen sich zwar biologische Geschlechter definieren, das sagt aber nicht ansatzweise etwas über Geschlechterrollen und gesellschaftliche Fragen etwas aus.
Crossing Over
Die Meiose und sexuelle Fortpflanzung ermöglicht des Weiteren die Erzeugung genetischer Vielfalt. Während die Mitose, salopp formuliert, eine Form des Klonens ist, sorgt die Meiose für genetische Rekombination.
Während der Prophase I und der Metaphase I setzt das Chromatin seine Spiralisierung und Komprimierung fort, sodass die Chromosomen immer dicker erscheinen. Zu einem bestimmten Zeitpunkt werden die homologen Chromosomen durch einen Spindelapparat aus Mikrotubuli voneinander weggezogen, vor allem in der Nähe der Centromere, aber sie bleiben über die Cohesine noch miteinander verbunden. Später in der Prophase I nehmen die Bereiche, in denen diese Anheftungen auftreten, die Form eines X an und man bezeichnet sie als Chiasmata. Ein Chiasma macht den Austausch von genetischem Material zwischen Nicht-Schwesterchromatiden auf homologen Chromosomen sichtbar – in der Genetik spricht man hier von einem Crossing-over (Abb. 5).
Ein Crossing-over führt zu rekombinanten (neu kombinierten) Chromatiden. Diese Rekombination des Erbmaterials erhöht die genetische Variabilität unter den Produkten der Meiose, indem genetische Information innerhalb der jeweiligen homologen Paare verschoben wird.
Das Crossing-over ist nur eine der Ursachen für die genetische Vielfalt der Meioseprodukte. Die andere Ursache ist die unabhängige Verteilung. Dabei erhält jede haploide Zelle aus der diploiden Zelle einen vollständigen Satz von Genen, aber nur jeweils ein Gen von jedem Genpaar von jedem Elternteil. Die Entscheidung, welches Chromosom aus einem homologen Paar in der Anaphase I in welche Tochterzelle gelangt, erfolgt rein zufällig. Stellt euch beispielsweise vor, in einem diploiden Zellkern seien zwei homologe Chromosomenpaare vorhanden.
Ein bestimmter Tochterzellkern könnte das väterliche Chromosom 1 und das mütterliche Chromosom 2 erhalten. Oder er könnte das väterliche Chromosom 2 und das mütterliche Chromosom 1 oder beide mütterlichen oder beide väterlichen Chromosomen erhalten. Das alles hängt davon ab, wie sich die homologen Paare in der Metaphase I zufällig anordnen. Je größer die Anzahl der Chromosomen, desto geringer ist die Wahrscheinlichkeit, dass die ursprünglichen elterlichen Kombinationen erneut entstehen, und umso größer ist das Potenzial für das Entstehen von genetischer Vielfalt.
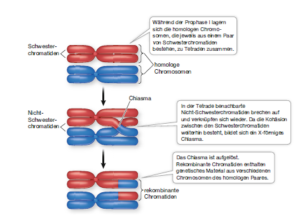
Abb. 5: Crossing over
Kernphasenwechsel
Dieser regelmäßige Wechsel zwischen einem durch die Meiose eingeleiteten haploiden Zustand (Haplophase) und einem aus der Befruchtung resultierenden diploiden Zustand (Diplophase) ist für alle Eukaryoten, die sich sexuell Vermehren, kennzeichnend. Organsimen haben dabei also immer einen Wechsel zwischen einer haploiden Phase (Gameten) und einer diploiden. Je nachdem welche der beiden Phasen überwiegt, kann man sie drei verschiedenen Typen zuordnen (Abb. 6)
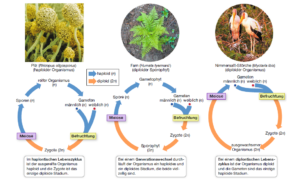
Abb. 6: Kernphasenwechsel
Haplonten
Bei Haplonten spielt sich fast der gesamte Entwicklungszyklus in der Haplophase (1n) ab. Nur die Zygote (= befruchtete Eizelle) besitzt einen diploiden Chromosomensatz (2n). Diese entsteht durch die Verschmelzung zweier Gameten. Die so entstandene Zygote vollzieht danach eine Reduktionsteilung (Meiose). So entstehen wieder 4 haploide Zellen (Gonen) und aus den haploiden Zellen kann durch Mitose wieder ein haploider Vielzeller entstehen. Zu den Haplonten gehören viele Pilze, viele Algen und einige tierische Einzeller, wie die Flagellaten.
Diplonten
Die Diplonten sind quasi das Gegenteil der Haplonten. Bei ihnen vollzieht sich die Lebensphase fast ausschließlich in der Diplophase. Nach der Befruchtung erfolgt, anders als bei den Haplonten, keine Reduktionsteilung (Meiose), sondern die Zygote entwickelt sich zum diploiden Organismus. Einige dieser Zellen bilden dann die haploiden Gameten, wodurch sich die Organismen wieder sexuell vermehren können. Neben einigen Algen und Pilzen gehören alle vielzelligen Tiere (Metazoa) und die Ciliata (Wimpertierchen, Einzeller) zu den Diplonten.
Diplo-Haplonten
Die Diplo-Haplonten (auch Haplo-Diplonten genannt) haben eine Zwischenstellung zu den zwei oben genannten Typen. Es sind Lebewesen, bei deren Fortpflanzung abwechselnd haploide und diploide Generationen auftreten. Hierzu gehören die meisten Pflanzen. Kennzeichnend für Diplo-Haplonten ist, dass sowohl in der haploiden wie in der diploiden Phase Mitosen erfolgen. Die haploide Generation pflanzt sich geschlechtlich fort, indem sie weibliche und männliche Gameten bildet. Sie wird deshalb als Gametophyt bezeichnet. Durch die Vereinigung zweier Gameten verschiedenen Geschlechts und Befruchtung entsteht eine diploide Zygote. Die Zygote wächst zum sogenannten Sporophyten. Die Sporophyten bilden nach einer Reduktionsteilung (Meiose) haploide Sporen. Aus diesen haploiden Sporen wächst der haploide Gametophyt heran und der Zyklus beginnt von vorne. In der Evolutionsgeschichte der Pflanzen kommt es dabei zur Reduktion des haploiden Gametophyten. Bei den Moosen ist der Gametophyt die dominante Phase (nämlich die grüne Moospflanze). Schon bei den Farnen ist der diploide Sporophyt dominant und bei den Samenpflanzen ist die eigentliche Pflanze der diploide Sporophyt, während der haploide Gametophyt auf wenige Zellen reduziert ist (männlicher Pollenschlauch, bestehend aus drei Zellen und weiblicher Embryosack in der Samenanlage, bestehend aus 7-8 Zellen).
