

In der letzten Episode haben wir kennengelernt, an welchen Merkmalen man einen Dinosaurier erkennt. Wenn ihr diese noch nicht kennt, schaut euch die entsprechende Episode nochmal an.
Dinosaurier sind Teil der Klade der Dinosauromorpha, die wiederum Teil der Klade der Ornithodira ist. Die Schwestergruppe der Dinosauromorpha sind die Pterosauromorpha, zu denen die Flugsaurier gehören. Ornithodira sind Teil der Avemetatarsalia und diese, zusammen mit den Pseudosuchia bzw. Crurotarsi, also der Krokodilverwandtschaft, Teil der Archosauria.
Text als pdf
Ursprünge der Dinosaurier
Wann traten die ersten Dinosaurier auf? Die Fossilien der ältesten unzweifelhaften Dinosaurier Eoraptor und Herrerasaurus entstammen der etwa 230 Millionen Jahre alten Ischigualasto-Formation in Argentinien, also aus der späten Trias (Abb. 1). Eoraptor gilt als der ursprünglichste Vertreter und sah wahrscheinlich dem gemeinsamen Vorfahren aller Dinosaurier sehr ähnlich (Sereno 1999, Sereno et al. 1988, 1993, 2013, Sereno & Novas 1992, Sterling et al. 2012, Reig 1963, Novas et al. 2021). Somit dürften die ersten Dinosaurier kleine, bipede Fleischfresser gewesen sein.
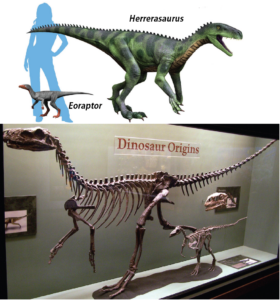
Abb. 1: Eoraptor und Herrerasaurus als die frühesten Dinosaurier
In Ostafrika wurde das Fossil Nyasasaurus entdeckt, der vor 245 Mio. Jahren lebte und der entweder als der früheste Dinosaurier oder als der nächste bekannte Verwandte der Dinosaurier in Frage kommt (Nesbitt et al. 2013, 2020, Novas et al. 2021). Wenn letzteres zutrifft, gehört er zu der Klade der Dinosauriformes innerhalb Dinosauromorpha, welches die Schwestergruppe der Dinosaurier wäre. Da nur wenige und fragmentierte Fossilien von Nyasasaurus gefunden wurden, lassen sich nur wenige morphologische Merkmale der Tiere feststellen, was seine genaue Einordnung im Stammbaum unsicher macht. Die Dinosauromorpha haben zwar einige Merkmale der „echten Dinosaurier“, aber nicht alle. Für den Nicht-Spezialisten sehen sie oberflächlich wie Dinosaurier aus, sind es aber noch nicht ganz. Einige kleinere Gattungen wie Lagosuchus, Marasuchus und Silesaurus gehören dazu (Angolin & Martin 2019, Dzik 2003, Sereno et al. 1994, Abb. 2). Aus solchen oder ähnlichen Formen gingen dann die Dinosaurier mit ihren eigenen Apomorphien hervor. Sind einige Dinosauromorpha den Dinosauriern ähnlicher als andere, aber immer noch keine Dinosaurier, sind sie einfach (zusammen mit den Dinosauriern) Dinosauriformes.
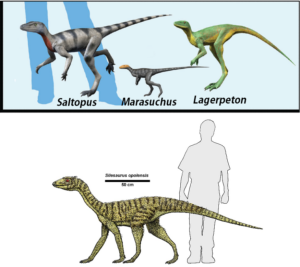
Abb. 2: Dinosauromorpha
Für das bessere Verständnis schauen wir uns einen Vertreter genauer an: Silesaurus. Er lebte vor 230 Mio. Jahren, war ca. 2m lang, hatte vier schlanke lange Beine und einen grazilen Körperbau. Äußerlich schon einem Dinosaurier sehr ähnlich, weist er dennoch einige entscheidende Unterschiede zu den „echten Dinosauriern“ auf. Seine Oberschenkelknochen verliefen zwar senkrecht, waren aber seitlich mit der Hüfte verbunden, also anders als bei Dinosauriern. Und weil der Gelenkkopf des Oberschenkelknochens bei Silesaurus weder geneigt noch in eine – für Dinosaurier typische – offene Gelenkpfanne eingepasst war und die Beine nicht unter dem Körper lagen, war er kein echter Dinosaurier. Obwohl Silesaurus auf vier Beinen lief, dienten die Vorderbeine eher zum Abstützen und nicht zur Fortbewegung. Es war nur noch ein kleiner Schritt, bis die ersten Dinosaurier ihren Körper so ausbalancierten, dass sie sich zweibeinig fortbewegen konnten. Das bot den Vorteil schneller laufen zu können. Gleichzeitig eröffnete es neue Anwendungsmöglichkeiten für die Hände, sie konnten z. B. nach der Nahrung greifen. Dies war das Erfolgsrezept der Dinosaurier und der Grund, weshalb sie sich gegen die anderen Dinosauromorpha zum Ende der Trias durchsetzen konnten (Schmitt 2023).
Saurischia – Ornithischia – Ornithoscelida
Wie verlief ausgehend von frühen Dinosauriern wie Eoraptor die Evolutionsgeschichte der Dinosaurier? Wir haben im letzten Beitrag kennengelernt, dass es zwei Hauptkladen innerhalb der Dinosaurier gibt: die Echsenbecken-Dinosaurier (Saurischia), welche die fleischfressenden Theropoden und die langhalsigen Sauropodomorpha vereinen und die Vogelbecken-Dinosaurier (Ornithischia), zu denen alle anderen pflanzenfressenden Dinosaurier wie Triceratops, Stegosaurus und Iguanodon gehören (Abb. 3). Trotz des Begriffs „Vogelbeckendinosaurier“ gehören Vögel nicht zu den Vogelbeckendinosaurier, sondern zu den Echsenbeckendinosaurier, da sie aus den Theropoden hervorgingen (Fastovsky & Weishampel 2021, Holtz 2007, Paul 2016, Prothero 2017, 2021, 2022, Prothero & Dott 2004, Brusatte 2019, Schweitzer et al. 2021).
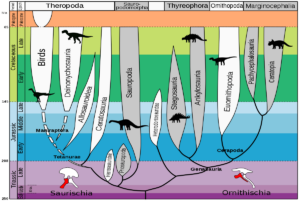
Abb. 3: Stammbaum der Dinosaurier
Die Ähnlichkeit zwischen dem Becken des Vogelbeckendinosaurier und der Vögel liegt in der Position des Schambeins. Anders als bei den Echsenbeckendinosauriern zeigt das Schambein (Pubis) bei den Vogelbeckendinosauriern nicht nach unten und leicht nach vorn zum Kopf, sondern besteht aus einem schmalen, stabförmigen Knochen, der längs neben dem Sitzbein (Ischium) liegt und nach hinten zeigt. Bei Vögeln ist das Schambein komplett nach hinten verlagert (Abb. 4).
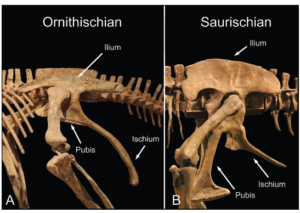
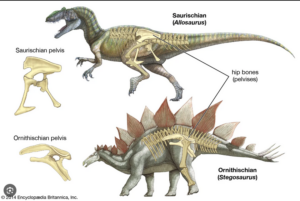
Abb. 4: Becken der Ornithischia und Saurischia
In einer detaillierten phylogenetischen Studie aus dem Jahr 2017, basierend auf 74 Taxa und 457 Merkmalen, schlagen die Paläontologen Matthew G. Baron, David B. Norman und Paul M. Barrett eine radikal neue Systematik der Dinosaurier vor (Baron et al. 2017a, Padian 2017). Demnach werden Ornithischia und Theropoda als Schwestergruppen in der Klade Ornithoscelida vereint, während die Sauropodomorpha zusammen mit den Herrerasauridae die neu definierte Klade Saurischia bilden (Abb. 5). Die Ornithoscelida-Hypothese wurde im November 2017 von einem internationalen Forscherteam in Frage gestellt, nachdem der ursprüngliche anatomische Datensatz von Baron et al. (2017a) überarbeitet worden war. Diese Überarbeitung ergab das traditionelle Modell, bei dem Ornithischia und Saurischia als Schwestertaxa wiedergefunden wurden. Dieser traditionelle Baum war jedoch nur schwach unterstützt und unterschied sich statistisch nicht signifikant von der alternativen Ornithoscelida-Hypothese (Langer et al. 2017). Mit nur geringfügigen Anpassungen, die Baron und Kollegen daraufhin vornahmen, wurde Ornithoscelida erneut gegenüber dem traditionellen Modell bevorzugt (Baron et al. 2017b).
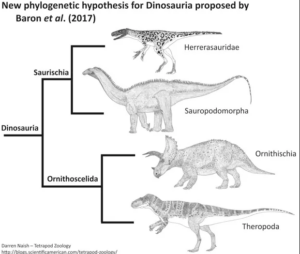
Abb. 5: Ornithoscelida-Konzept
Weil die Debatte um die Validität des Ornithoscelida-Konzeptes noch nicht abgeschlossen ist und die meisten Dinosaurier-Lehrbücher nach dem traditionellen Konzept gehen, bleiben wir auch bei diesen. Die Verwandtschaftsverhältnisse innerhalb der Ornithischia, Theropoden und Sauropodomorpha sind aber auch beim Ornithoscelida-Konzept mit dem traditionellen Konzept vereinbar.
Wenn man bedenkt, wie selten Dinosaurierfossilien vor allem im Vergleich zu den wirbellosen Meerestieren sind, ist es bemerkenswert, dass wir nicht nur eine recht große Vielfalt an Dinosauriergattungen haben – etwa 1000 beschriebene Gattungen – sondern auch genügend Exemplare haben, um zu zeigen, dass wir Übergänge zwischen fast allen großen Dinosauriergruppen zeigen, sowie viele andere bemerkenswerte Fossilien, die andere Arten von Übergängen zeigen, wie z. B. fleischfressende Dinosaurier, die zu Pflanzenfressern wurden. In diesem Beitrag wollen wir uns zuerst mit jenen Dinosauriern befassen, die nichts mit Vögeln zu tun haben.
Evolutionäre Übergänge bei den Sauropodomorpha
Zu den atemberaubendsten Dinosauriern gehören die gigantischen und langhalsigen Sauropoden. Unter ihnen fanden sich nicht nur Giganten wie Diplodocus, Brachiosaurus oder Argentinosaurus, sondern auch Zwergformen wie Europasaurus (Abb. 6). Die Ursache für den Gigantismus der Sauropoden wurde in einem separaten Beitrag von mir schon behandelt[1] (Sander & Clauss 2008, Sander et al. 2011, 2013, Klein et al. 2013, Clauss et al. 2013, Christian et al. 2013, Stevens 2013, Taylor & Wedel 2013a,b, Sellers et al. 2013, Wedel & Taylor 2013, Fastovsky & Weishampel 2021, Holtz 2007, Paul 2016, Prothero 2022, Schweitzer et al. 2021, Manz 2022). Spannender für uns ist: gibt es hier eigentlich evolutionäre Übergänge? Ja!

Abb. 6: Größenvergleich der Sauropoden Argentinosaurus und Europasaurus
In fast jedem Buch über Dinosaurier (z. B. Fastovsky & Weishampel 2021, Holtz 2007, Paul 2016, Schweitzer et al. 2021) wird eine Gruppe von Kreaturen aus der Trias abgebildet, die Prosauropoden genannt werden. Schon ihr Name (übersetzt „vor den Sauropoden“) deutet darauf hin, dass sie ursprüngliche Verwandte der größeren Sauropoden sind. Der bekannteste von ihnen ist Plateosaurus aus der Trias in Deutschland, aber es gibt noch ein Dutzend weitere Gattungen, die in Trias-Schichten auf der ganzen Welt gefunden wurden (Abb. 7 – 9). Die meisten dieser Gattungen waren nur etwa 5-8 Meter lang, also nur ein Viertel so groß wie die riesigen jurassischen Sauropoden, aber größer als ihre Vorfahren. Sie haben die Anfänge eines langen Halses und eines langen Schwanzes, aber noch nicht die unglaublichen Ausmaße der Riesensauropoden. Die Gliedmaßen sind in der Konstruktion der Finger und Zehen klassisch sauropodisch, aber nicht so robust, und die Vordergliedmaßen sind lang und zart genug, dass sie zwischen dem vierfüßigen Gehen und dem Aufrichten auf die Hinterbeine in einer zweibeinigen Haltung wechseln konnten, um ihre Hände zu benutzen. Erst als die Sauropoden eine enorme Größe erreichten, waren sie gezwungen, ausschließlich auf allen Vieren zu gehen, und auch ihre Gliedmaßen wurden viel massiver, um ihr enormes Körpergewicht zu tragen.

Abb. 7: Plateosaurus und Massospondylus
Abb. 8: verschiedene Prosauropoden: A) Guaibasaurus, (B) Panphagia, (C) Saturnalia, (D) Pantydraco, (E) Bagualosaurus, (F) Thecodontosaurus
Abb. 9: der Prosauropode Plateosaurus im Vergleich zu den Sauropoden (Brachiosaurus und Barosaurus). Wie andere basalen Sauropodomorpha, hatte Plateosaurus hatte einen verlängerten Hals und war zweibeinig. Alle Sauropodomorpha haben mindestens 10 Halswirbel (bei Plateosaurus ist Halswirbel Nr. 1 in der Abbildung oben links nicht zu sehen). Die Zahl der Halswirbel hat sich bei Sauropoden im Verlauf der Evolution erhöht, Barosaurus hatte 16. Beim Gigantismus kam es im Verlauf der Evolution der Sauropodomorpha zu einer Umorganisation der Vordergliedmaßen (dargestellt am Beispiel des Brachiosaurus, rechts im Bild). Die Oberarmknochen, Speiche und Elle sowie die Mittelhandknochen sind in einer vertikalen Säule angeordnet. Außerdem sind die Handwurzelknochen und Finger reduziert worden. Zu sehen ist auch ein Sauropoden-Rückenwirbel mit beschrifteter Lamina und Fossa. Diese Strukturen sind das Ergebnis der postkranialen Pneumatisierung, einem Prozess, bei dem Erweiterungen der Lunge in den Knochen eindringen. Entwicklung.


Noch ursprünglicher als Plateosaurus war Anchisaurus aus der Trias von Connecticut, Arizona und Südafrika (Yates 2004, Prothero 2017, Abb. 10). Er war nur 2,5 Meter lang, also nur wenig größer als ein Mensch, und hatte einen noch kürzeren Hals und Schwanz sowie noch zartere Gliedmaßen und Füße. In der Tat ist er die perfekte Übergangsform zwischen den echsenartigen frühen Sauriern wie dem viel kleineren Lagosuchus und größeren Formen wie Plateosaurus. Trotz seiner äußeren Erscheinung trägt sein Schädel alle charakteristischen Merkmale der Sauropoden, und er zeigt bereits viele der Spezialisierungen an den Wirbeln und vor allem an den Händen und Füßen, die später die Sauropoden kennzeichnen werden. Die frühen Saurier waren in erster Linie zweibeinig, aber Anchisaurus scheint zu beiden Haltungen fähig gewesen zu sein, und Plateosaurus war noch schwerer und eher vierbeinig.

Abb. 10: Anchisaurus
Vom frühesten bekannten Dinosaurier Eoraptor über Anchisaurus und Plateosaurus bis hin zu den größeren Sauropoden gibt es nicht nur einen fließenden Größenzuwachs, sondern auch einen fließenden Übergang in den anatomischen Merkmalen und in der Köperhaltung vom Zweibeiner zum Vierbeiner (Abb. 11).
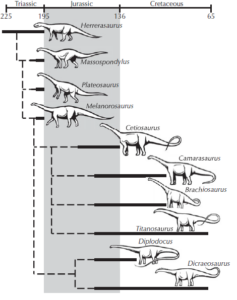
Abb. 11: Stammbaum der Sauropoden. Die Prosauropoden waren Übergangsformen zwischen kleineren primitiven zweibeinigen Dinosauriern wie Eoraptor und Herrerasaurus und den Sauropoden.
Evolutionäre Übergänge bei den Nicht-Vogel-Theropoden
Der andere Hauptzweig der Echsenbeckendinosaurier waren die Theropoden, also die Raubsaurier, zu denen der berühmte Tyrannosaurus rex gehört, aber es gibt Dutzende verschiedener Gattungen und Arten in allen Formen und Größen. Die frühen Theropoden waren jedoch recht unscheinbar und sind tatsächlich hervorragende Übergangsformen. Ihre Länge reichte von nur 70 Zentimetern (Compsognathus) bis zu 3 Metern (Coelophysis), d. h. die meisten waren etwa so groß wie ein Huhn bis hin zur Größe eines erwachsenen Menschen. Anders als ihre großen Theropoden-Nachfahren waren sie leicht gebaut, mit kleinen Köpfen, langen Hälsen und schlanken, grazilen Gliedmaßen und Schwänzen. Dennoch wiesen ihre Schädel und vor allem ihre Hände (mit ihrer einzigartigen Kombination aus nur drei Fingern: Daumen, Zeige- und Mittelfinger) und Füße alle anatomischen Spezialisierungen auf, die man bei späteren Theropoden findet.
Von ursprünglichen Theropoden wie Coelophysis (Abb. 12) können wir die Theropodenlinie noch weiter zurückverfolgen bis zu Eoraptor, Staurikosaurus und Herrerasaurus, die ähnlich gebaut sind wie Coelophysis, aber noch nicht alle ausgeprägten Spezialisierungen der Theropoden aufweisen (Fastovsky & Weishampel 2021, Holtz 2007, Paul 2016, Prothero 2017, 2021, 2022, Prothero & Dott 2004, Brusette 2012, 2019, Schweitzer et al. 2021).
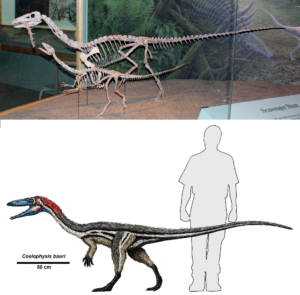
Abb. 12: Coelophysis
Für einen zufälligen Beobachter sehen sie sehr ähnlich aus, aber für einen Paläontologen mit anatomischer Ausbildung sind die Unterschiede deutlich. Eoraptor, Staurikosaurus und Herrerasaurus fehlt die hochspezialisierte dreifingrige Hand (einige hatten noch die vollen fünf Finger, Abb. 13), die relativ unspezialisierten Wirbel (sowohl Sauropoden- als auch Theropodenwirbel sind sehr spezialisiert und ausgeprägt), ein gleitendes Kiefergelenk, die vollständig gebogenen Raubtierzähne und die für Theropoden typischen Modifikationen an Knöchel und Fuß, die man bei Coelophysis und fortgeschritteneren Theropoden findet (Schwartz et al. 1994, Rinehart et al. 2004, 2007, 2009, Gay 2001, Downs 2000, Barta et al. 2018, Prothero 2017, Ezcurra 2007) Schließlich können wir Gattungen wie Eoraptor, Staurikosaurus und Herrerasaurus auf noch ursprünglichere, nicht dinosaurierähnliche Archosaurier wie Euparkeria zurückführen, die oberflächlich betrachtet den frühesten Dinosauriern ähneln, denen aber die einzigartigen Spezialisierungen fehlen, die alle Dinosaurier haben, wie z. B. die offene Hüftgelenkpfanne oder die charakteristischen Merkmale des Schädels.
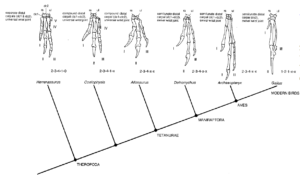
Abb. 13: Die linke Hand ausgewählter Theropoden im phylogenetischen Kontext zeigt das Muster der Finger-Reduktion und die Entwicklung des Handgelenks vom Universalgelenk zum Drehgelenk. Die Problematik der Zählweise der Finger bei Theropoden (I-II-III) und Vögel (II-III-IV) wird in einem anderen Beitrag gesondert besprochen.
So können wir sowohl die Sauropoden (über die Prosauropoden) als auch die Theropoden (über die primitiven Gattungen wie Coelophysis) auf einen gemeinsamen Saurischia-Vorfahren in der Art von Eoraptor, Staurikosaurus und Herrerasaurus zurückführen, und von dort zu primitiveren Archosauriern, die keine Dinosaurier waren, wie Euparkeria. Eine schönere Reihe von Übergangsformen kann man sich nicht wünschen. Offensichtlich haben Kreationisten noch nie von einer dieser Formen gehört. Und es gibt noch eine coole letzte Wendung in dieser Geschichte der Raubsaurier. Im Jahr 2005 gaben Jim Kirkland und andere die Entdeckung eines bemerkenswerten neuen Fossils mit dem Namen Falcarius utahensis aus dem Jura von Utah bekannt (Kirkland et al. 2005, Zanno 2010a,b, Zanno & Erickson 2006, Abb. 14). Diese seltsame Kreatur gehört zu einer noch seltsameren Gruppe, den Therizinosauriern, deren genaue Position innerhalb der Dinosaurier lange umstritten war. Diese Tiere haben viele der Merkmale von Theropoden wie Velociraptor, einschließlich langer Krallenfinger an den Händen und eines langen Halses und Schwanzes zum Balancieren. Aber sie hatten zahnlose Schnäbel und waren offenbar Pflanzenfresser. In den letzten Jahren herrschte Einigkeit darüber, dass es sich bei den Therizinosauriern tatsächlich um Theropoden handelte, die irgendwie zum Pflanzenfressen zurückgekehrt waren. Die Entdeckung von Falcarius lieferte das „fehlende Glied“ in dieser Ernährungsumstellung, da er viele Merkmale von „Raptoren“-Dinosauriern beibehält, jedoch der primitivste Therizinosaurier mit einem zahnlosen pflanzenfressenden Schnabel ist. Damit war er eine klassische Übergangsform, die die Therizinosaurier nicht nur anatomisch mit den Raptoren verband, sondern auch zeigte, wie sie den bemerkenswerten Übergang vom Fleischfresser zurück zum Raptor vollzogen.

Abb. 14: Falcarius utahensis
Evolutionäre Übergänge bei den Ornithischia
Der andere Hauptzweig der Dinosaurier sind die Ornithischia, zu der fast alle pflanzenfressenden Dinosaurier (mit Ausnahme der Sauropoden) gehören: die Entenschnäbel, die Iguanodonten, die gepanzerten Ankylosaurier, die stacheligen Stegosaurier, die dickköpfigen Pachycephalosaurier und die gehörnten Ceratopsier. Zu den frühesten Ornithischiern gehören primitive triassische Formen wie Lesothosaurus, Fabrosaurus und Heterodontosaurus, kleine zweibeinige Dinosaurier, die oberflächlich betrachtet wie Eoraptor oder Coelophysis aussehen (Abb. 15-17). Bei näherer Betrachtung weisen sie jedoch alle Merkmale der Ornithischia auf: ein Teil oder das gesamte Schambein in der Hüfte ist parallel zum Sitzbein nach hinten gedreht; die Backenzähne sind tief in den Kiefer eingesetzt, was darauf hindeutet, dass sie Wangen hatten, um ihre Nahrung beim Kauen im Mund zu halten; und sie haben einen einzigartigen zusätzlichen Knochen an der Spitze des Unterkiefers, der als Praedentale bekannt ist (Abb. 18). All diese Merkmale sind einzigartig für die Ornithischia, doch wir können sehen, dass die Übergangsformen wie der Heterodontosaurus sie bereits in der Trias hatten, während sie noch den anderen primitiven Dinosauriern dieser Zeit ähnelten (Fastovsky & Weishampel 2021, Holtz 2007, Paul 2016, Prothero 2017, 2022, Schweitzer et al. 2021).
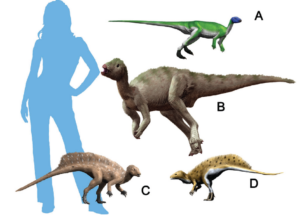
Abb. 15: ursprüngliche Ornithischia. (A) Lesothosaurus, (B) Hypsilophodon, (C) Heterodontosaurus, (D) Tianyulong.
Abb. 16: frühe Ornithischia. (A) Hypsilophodon, (B) Leaellynasaura, (C) Orodromeus, (D) Kulindadromeus.
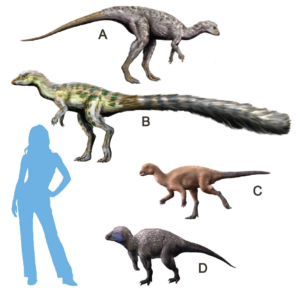

Abb. 17: Skelett des Heterodontosaurus

Abb. 18: Synapomorphien der Ornithischia. Links: Schambein eines Lambeosaurus (Abguss). Bei diesem und anderen Ornithischia zeigt das Schambein nach hinten (Pfeile zeigen das kaudale Ende des Dinosauriers, fast parallel zum Sitzbein. Rechts: Schädel (Abguss) eines Maiasaura, mit markierten ornithischen Merkmalen, einschließlich des Praedentale und eines Kiefergelenks, das unterhalb der Zahnreihe des Oberkiefers liegt.
Zu den beliebtesten Ornithischia gehört Triceratops, einer der letzten gehörnten Dinosaurier oder Ceratopsier. Die Ceratopsier sind ein weiterer klassischer Fall von Übergangsformen zwischen hochspezialisierten Formen, wie den gehörnten und gekräuselten Ceratopsiern, und viel primitiveren Formen, die dem gemeinsamen Vorfahren mit anderen Dinosaurierstämmen ähneln (Abb. 19 und 20).

Abb. 19: gehörnte Ceratopsia. (E) Diabloceratops, (F) Pachyrhinosaurus, (G) Styracosaurus, (H) Agujaceratops, (I) Chasmosaurus, (J) Pentaceratops, (K) Triceratops

Abb. 20: frühe Ceratopsia. (A) Yinlong, (B) Psittacosaurus, (C) Protoceratops, (D) Leptoceratops
Alle Gattungen mit Hörnern lassen sich auf den sehr bekannten Protoceratops zurückführen, der die Halskrause und die charakteristischen Knochen im Schnabel und Unterkiefer hat, dem aber die Hörner fehlen (Fastovsky & Weishampel 2021, Holtz 2007, Paul 2016, Weishampel et al. 1990, Granger & Gregory 1923, Abb. 20 & 21). Kreationisten, wie der US-Guru Duane Gish scheinen aber die Stellung von Protoceratops als Übergang zwischen den Ceratopsiern nicht zu verstehen (Gish 1995). Er zitiert ein aus dem Zusammenhang gerissenes Zitat aus der ersten Ausgabe von Weishampel et al. (1990) über die Besonderheit der Protoceratopsidae (und übersieht dabei völlig, dass dies sie nicht weniger zu einer guten Übergangsform macht), und er erwähnt auch, dass sie in der späten Kreidezeit vorkommen und daher keine Vorfahren sein können. Ich habe schon in meinem Beitrag über das Thema der Übergangsformen erwähnt, dass Paläontologen nicht nach Vorfahren suchen, sondern nach Schwestergruppen, die anatomische Übergänge belegen. Aufgrund der Seltenheit der Fossilisation ist nicht zu erwarten, dass man den direkten Vorfahren einer Evolutionslinie entdeckt. Zweitens kam Protoceratops in der frühen Spätkreide vor, Millionen von Jahren vor all seinen mutmaßlichen Nachfahren unter den gehörnten Ceratopsia in der späten Spätkreide. Außerdem wir haben noch bessere Übergangsformen.

Abb. 21: Protoceratops
Bagaceratops hat im Vergleich zu Protoceratops eine etwas kleinere Halskrause und einen kleineren Schnabel, und sein Körper ist nicht vollständig vierfüßig (Maryanska & Osmolska 1975, Czepinsky 2019, Abb. 22). Archaeoceratops hat eine noch kleinere Halskrause und einen noch kleineren Schnabel und einen viel leichteren Körper, der höchstwahrscheinlich zweibeinig ist (Tang et al. 2001, Jin et al. 2009, Abb. 23). Psittacosaurus, der so genannt wurde, weil er einen papageienähnlichen Schnabel und die Anfänge eines Kragens über dem Hals hatte, ist viel leichter gebaut mit einem zweibeinigen, grazilen Skelett anstelle des schwereren Skeletts von Protoceratops. Psittacosaurus zeigt nicht nur den Übergang von einem kragenlosen Schädel zu einem mit einem kleinen Kragen zu dem größeren Kragen des Protoceratops, sondern auch den Übergang von einem leichten zweibeinigen Körper (typisch für fast alle ursprünglichen Dinosaurier) zu dem schwereren vierbeinigen Körper der spezialisierteren Gruppen (Osborn 1923, Coombs 1982, Sereno 2010, Mayr et al. 2002, Dalton 2001, Lingham-Soliar 2001, Vinther et al. 2016, 2021, Abb. 20 & 24).
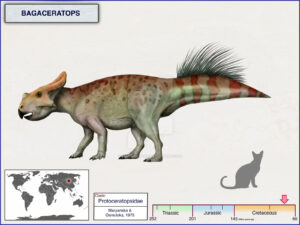
Abb. 22: Bagaceratops
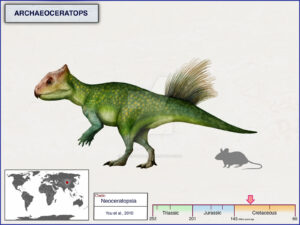
Abb. 23: Archaeoceratops

Abb. 24: Psittacosaurus.
Das erstaunlichste Übergangsfossil in dieser Abfolge wurde schließlich 2004 mit der Entdeckung von Yinlong aus den viel früheren Schichten des späten Juras in China entdeckt (Xu et al. 2006, Abb. 20 & 25). Der Name Yinlong bedeutet in Mandarin „verborgener Drache“, in Anlehnung an den populären Film „Crouching Tiger, Hidden Dragon“, der teilweise in der Nähe des Fundortes gedreht wurde. Yinlong besteht aus einem gut erhaltenen Skelett eines zweibeinigen Dinosauriers, der sich in seinen Proportionen nicht allzu sehr von Psittacosaurus unterscheidet. An der Spitze seines Oberschnabels hat er den für Ceratopsier typischen Rostralknochen. Sein Schädeldach weist jedoch eine einzigartige Anordnung von Knochen auf, wie man sie bei den dickschädeligen Pachycephalosauriern findet, die dafür bekannt sind, dass sie eine dicke Knochenkuppel in ihrem Schädel haben. Paläontologen haben lange Zeit argumentiert, dass Ceratopsia und Pachycephalosaurier eng miteinander verwandt sind, da sie beide einen Knochenkamm am hinteren Rand des Schädels haben. Beide werden als „Marginocephala“ (Fastovsky & Weishampel 2021, Holtz 2007, Paul 2016, Prothero 2017, 2022, Schweitzer et al. 2021) zusammengefasst (Abb. 26). Aber mit Yinlong haben wir ein wunderschönes Übergangsfossil, das Merkmale sowohl der Ceratopsier als auch der Pachycephalosaurier zeigt, bevor sich beide Familien aufspalteten.
Abb. 25: Fossil von Yinlong
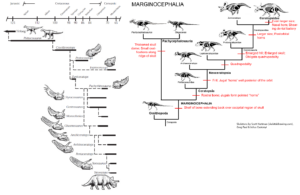
Abb. 26. Evolution der Marginocephalia
Erwähnenswert seien auch die Übergänge der gepanzerten Anklyosaurier und Stegosaurier. Beide Familien werden als Thyreophora zusammengefasst und zeichnen sich durch für diese Gruppe einzigartige Apomorphien aus, wie entlang des Körpers aufgereihte Panzerplatten, relativ kleine Gehirne und eine vierfüßige Haltung mit Hintergliedmaßen, die länger als die Vordergliedmaßen sind (Fastovsky & Weishampel 2021, Holtz 2007, Paul 2016, Prothero 2017, 2022, Schweitzer et al. 2021, Abb. 27).
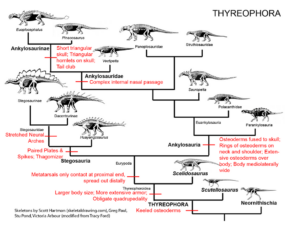
Abb. 27: Thyreophora
So haben die ursprünglichen Anklyosaurier – die Nodosaurier – noch eine sehr begrenzte Panzerung, genauso wie die ursprünglichen Stegosaurier, wie Scelidosaurus (Abb. 28). Wir können all diese Formen auf Gattungen wie Scutellosaurus zurückführen, einem kleinen, etwa 1,2 m langen Dinosaurier aus dem Unterjura vor 200 Mio. Jahren (Abb. 29). Er hatte noch längere Hinterbeine und konnte sich sowohl auf zwei als auch auf vier Beinen fortbewegen. Sein Körper war von kleinen Hornschuppen bedeckt, die vermutlich in Reihen am Rücken und an den Flanken des Tieres angebracht waren (Norman 2020, 2021, Norman et al. 2004 Gierlinski 1999, Maidment & Porro 2010, Martill et al. 2000, Carpenter 2001, Thompson et al. 2011, Colbert 1981, Prothero 2017, 2021, Fastovsky & Wieshampel 2021, Schweitzer et al. 2021).
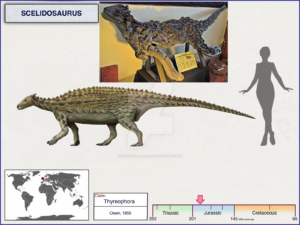
Abb. 28: Scelidosaurus
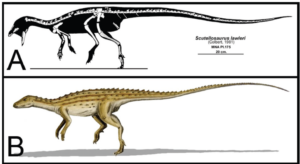
Abb. 29: Scutellosaurus
Alle Ornithischia können wir bis zu den ursprünglichsten Formen, wie Heterodontosaurus, Fabrosaurus oder Lesothosaurus zurückverfolgen. Diese Gattungen wiederum sahen in ihren äußeren Merkmalen Eoraptor und Herrerasaurus, den frühesten Dinosauriern, sehr ähnlich, abgesehen von ein paar subtilen Unterschieden, wie dem Vorhandensein eines Praedentale an der Spitze des Unterkiefers und einer primitiven ornithischen Hüftstruktur. Wenn man die Literatur sorgfältig und unvoreingenommen liest, sind diese Übergänge offensichtlich.
Wir könnten endlos fortfahren, kreationistische Unwahrheiten über Dinosaurier zu entlarven. Jeder, der nur ein mäßiges Interesse an diesem Thema hat, braucht sich nur ein Dinosaurierbuch zu kaufen – manchmal reicht schon ein Kinderbuch! – und sich die wunderschönen Übergangsformen für fast jede Gruppe ansehen, die Kreationisten abstreiten. Doch bis hier sollte es erstmal genügen. Wenn wir schon zwischen solchen verschiedenen Dinosauriern Übergänge finden, so ist der Übergang zwischen Vögeln und theropoden Dinosauriern sogar noch spannender. Und diesen lernen wir in der nächsten Episode kennen.
Literatur
Agnolin, F. L., Ezcurra, M. D. (2019): The Validity of Lagosuchus Talampayensis Romer, 1971 (Archosauria, Dinosauriformes), from the Late Triassic of Argentina. Breviora, 565(1):1-21
Baron, M. G., Norman, D. B., Barrett, P. M. (2017a): A new hypothesis of dinosaur relationships and early dinosaur evolution. Nature 543:501–506.
Baron, M. G., Norman, D. B., Barrett, P. M. (2017b). Baron et al. reply. Nature 551(7678): E4–E5.
Barta, D. E., Nesbitt, S. J., Norell, M. A. (2018): The evolution of the manus of early theropod dinosaurs is characterized by high inter‐ and intraspecific variation. Journal of Anatomy. 232 (1): 80–104.
Brusatte, S. L. (2019): The Rise and Fall of the Dinosaurs: A New History of a Lost World. William Morrow, NY
Carpenter, K. (2001): Phylogenetic Analysis of Ankylosauria. In Carpenter, Kenneth (ed.). The Armored Dinosaurs. Indiana University Press. pp. 455–480.
Clauss, M., Steuer, P., Müller, D. W. H., Codron, D., Hummel, J. (2013): Herbivory and Body Size: Allometries of Diet Quality and Gastrointestinal Physiology, and Implications for Herbivore Ecology and Dinosaur Gigantism. PLoS ONE 8(10): e68714.
Christian, A., Peng, G., Sekiya, T., Ye, Y., Wulf, M. G., Steuer, T. (2013): Biomechanical Reconstructions and Selective Advantages of Neck Poses and Feeding Strategies of Sauropods with the Example of Mamenchisaurus youngi. PLoS ONE 8(10): e71172.
Colbert, E. H. (1981): A primitive ornithischian dinosaur from the Kayenta Formation of Arizona. Museum of Northern Arizona Bulletin 53:1-61
Coombs, W. P. (1982): Juvenile specimens of the ornithischian dinosaur Psittacosaurus. Palaeontology. 25: 89–107
Czepiński, Ł. (2019): Ontogeny and variation of a protoceratopsid dinosaur Bagaceratops rozhdestvenskyi from the Late Cretaceous of the Gobi Desert. Historical Biology. 32 (10): 1394–1421.
Dalton, R. (2001): Wandering Chinese fossil turns up at museum. Nature 414: 571.
Downs, A. (2000): Coelophysis bauri and Syntarsus rhodesiensis compared, with comments on the preparation and preservation of fossils from the Ghost Ranch Coelophysis quarry: In: Lucas, S.G., Heckert, A.B. (eds.). „Dinosaurs of New Mexico“. New Mexico Museum of Natural History Bulletin. 17: 33–37
Dzik, J. (2003): A beaked herbivorous archosaur with dinosaur affinities from the early Late Triassic of Poland. Journal of Vertebrate Paleontology. 23 (3): 556–574.
Ezcurra, M. D. (2007): The cranial anatomy of the coelophysoid theropod Zupaysaurus rougieri from the Upper Triassic of Argentina. Historical Biology. 19 (2): 185–202.
Fastovsky, D. E., Weishampel, D. B. (2021): Dinosaurs: A Concise Natural History (4th ed.). Cambridge university Press, Cambridge.
Gay, R.J. (2001): An unusual adaptation in the caudal vertebrae of Coelophysis bauri (Dinosauria: Theropoda). PaleoBios. 21: 55.
Gierliński, G. (1999): Tracks of a large thyreophoran from the Early Jurassic of Poland. Acta Palaeontologica Polonica. 44.
Gish, D. (1995): Evolution, the Fossils Still Say NO! San Diego, Calif.: Creation-Life.
Granger, W. W., Gregory, W. K. (1923): Protoceratops andrewsi, a pre-ceratopsian dinosaur from Mongolia. American Museum of Natural History Novitates (72): 1−9.
Holtz, T. R. (2007): Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages
Jin L., Chen J., Zan S., Godefroit, P. (2009): A New Basal Neoceratopsian Dinosaur from the Middle Cretaceous of Jilin Province, China. Acta Geologica Sinica. 83 (2): 200–206.
Kirkland, J. I., Zanno, L. E., Sampson, S. D., Clark, J. M., DeBlieux, D. D. (2005): A primitive therizinosauroid dinosaur from the Early Cretaceous of Utah. Nature. 435 (7038): 84–87.
Klein, N., Remes, K., Gee, C. T., Sander, P. M. (2011): Biology of the Sauropod Dinosaurs: Understanding the Life of Giants. Bloomington: Indiana University Press
Langer, M. C., Ezcurra, M. D., Rauhut, O. W. M., Benton, M. J., Knoll, F., McPhee, B. W., Novas, F. E., Pol, D., Brusatte, S. L. (2017): Untangling the dinosaur family tree. Nature551(7678): E1–E3.
Lingham-Soliar, T. (2008): A unique cross section through the skin of the dinosaur Psittacosaurus from China showing a complex fibre architecture. Proceedings of the Royal Society B: Biological Sciences. 275 (1636): 775–780.
Maidment, S. C. R., Porro, L.B. (2010): Homology of the palpebral and origin of the supraorbital ossifications in ornithischian dinosaurs. Lethaia, 43: 95-111
Manz, A. (2022): Sauropoden – dem Gigantismus auf der Spur https://www.scinexx.de/dossier/sauropoden-dem-gigantismus-auf-der-spur/
Martill, D. M., Batten, D. J., Loydell, D. K. (2000): A New Specimen of the Thyreophoran Dinosaur cf. Scelidosaurus with Soft Tissue Preservation. Palaeontology 43(3): 549-559.
Maryańska, T., Osmólska, H. (1975): Protoceratopsidae (Dinosauria) of Asia. Palaeontologia Polonica. 33: 134−143.
Mayr, G., Peters, S. D., Plodowski, G., Vogel, O. (2002): Bristle-like integumentary structures at the tail of the horned dinosaur Psittacosaurus. Naturwissenschaften. 89 (8): 361–365.
Nesbitt, S. J., Barrett, P. M., Werning, S., Sidor, C. A., Charig, A. J. (2013): The oldest dinosaur? A Middle Triassic dinosauriform from Tanzania. Biol. Lett. 9 (1): 20120949.
Nesbitt, S, Butler, R, Ezcurra, M, Charig, A, Barrett, P (2020): The anatomy of Teleocrater Rhadinus, an early avemetatarsalian from the lower portion of the Lifua Member of the Manda Beds (Middle Triassic) (project). MorphoBank datasets.
Norman, D. B. (2020): Scelidosaurus harrisonii from the Early Jurassic of Dorset, England: cranial anatomy. Zoological Journal of the Linnean Society. 188 (1): 1–81.
Norman, D. B. (2021): Scelidosaurus harrisonii (Dinosauria: Ornithischia) from the Early Jurassic of Dorset, England: biology and phylogenetic relationships. Zoological Journal of the Linnean Society. 191(1):1–86.
Norman, D. B., Witmer, L. M., Weishampel, D. B. (2004): Basal Thyreophora. In Weishampel, D.B., Dodson, P., and Osmólska, H. (ed.). The Dinosauria, 2nd Edition. University of Californian Press. pp. 335–342
Novas, F. E., Agnolin, F. L., Ezcurra, M. D., Temp Müller, R., Martinelli, A. G., Langer, M. C. (2021): Review of the fossil record of early dinosaurs from South America, and its phylogenetic implications. Journal of South American Earth Sciences. 110: 103341.
Osborn, H. F. (1923): Two Lower Cretaceous dinosaurs of Mongolia. American Museum Novitates (13): 1–10.
Padian, K. (2017): Palaeontology: Dividing the dinosaurs. Nature 543:494–495.
Paul, G. S. (2016): The Princeton Fieldguide to Dinosaurs. 2nd Edition. Princeton University Press
Prothero, D. (2017): Evolution – What the Fossils say and why it matters. Second edition. New York: Columbia University Press
Prothero, D. (2021): The Evolving Earth. Oxford University Press
Prothero, D. (2022): Vertebrate Evolution. From Origins to Dinosaurs and beyond. CRC Press, Kapitel 11, 12, 14
Prothero, D. & Dott, (2004): Evolution of the Earth, Seventh edition. McGrawHill
Reig, O.A. (1963): La presencia de dinosaurios saurisquios en los „Estratos de Ischigualasto“ (Mesotriásico Superior) de las provincias de San Juan y La Rioja (República Argentina). Ameghiniana 3 (1): 3–20.
Rinehart, L.F., Heckert, A.B., Lucas, S.G., Hunt, A.P. (2004): The sclerotic ring of the Late Triassic theropod dinosaur Coelophysis. New Mexico Geological Society Spring Meeting. 26: 64.
Rinehart, L.F., Lucas, S.G., Heckert, A.B., Spielmann, J.A., Celesky, M.D. (2009): The paleobiology of Coelophysis bauri (Cope) from the Upper Triassic (Apachean) Whitaker quarry, New Mexico, with detailed analysis of a single quarry block. New Mexico Museum of Natural History & Science, A Division of the Department of Cultural Affairs Bulletin. 45: 260.
Rinehart, L. F., Lucas, S. G., Hunt, A. P. (2007): Furculae in the Late Triassic theropod dinosaur Coelophysis bauri. Paläontologische Zeitschrift. 81 (2): 174–180.
Sander, P. M. (2013): An Evolutionary Cascade Model for Sauropod Dinosaur Gigantism – Overview, Update and Tests. PLoS ONE 8(10): e78573.
Sander, P. M., Clauss, M. (2008): Sauropod gigantism. Science 322: 200–201.
Sander, P. M, Christian, A., Clauss, M., Fechner, R., Gee, C. T., et al. (2011): Biology of the sauropod dinosaurs: the evolution of gigantism. Biological Reviews 86: 117–155.
Schmitt, A. (2023): Großartige Giganten. Den letzten Geheimnissen der Dinosaurier auf der Spur. Dtv Verlagsgesellschaft
Schwartz, H. L., Gillette, D. D. (1994): Geology and taphonomy of the Coelophysis quarry, Upper Triassic Chinle Formation, Ghost Ranch, New Mexico. Journal of Paleontology. 68 (5): 1118–1130.
Schweitzer, M. H., Schroeter, E. R., Czajka, C. D. (2021): Dinosaurs How We KNow What We Know. CRC Press
Sellers, W. I., Margetts, L., Coria, R. A., Manning, P. L. (2013): March of the Titans: The Locomotor Capabilities of Sauropod Dinosaurs. PLoS ONE 8(10): e78733.
Sereno, P. C. (1999): The Evolution of Dinosaurs. Science 284(5423): 2137–2147
Sereno, P.C. (2010): Taxonomy, cranial morphology, and relationships of parrot-beaked dinosaurs (Ceratopsia:Psittacosaurus). In Ryan, Michael J., Chinnery-Allgeier, Brenda J., Eberth, David A. (eds.). New Perspectives on Horned Dinosaurs: The Royal Tyrrell Museum Ceratopsian Symposium. Bloomington and Indianapolis: Indiana University Press. pp. 21–58.
Sereno, P. C., Arcucci, A. B. (1994): Dinosaurian precursors from the Middle Triassic of Argentina: Marasuchus lilloensis, gen. nov. Journal of Vertebrate Paleontology. 14 (1): 53–73.
Sereno, P.C., Forster, C.A., Rogers, R.R., Moneta, A.M. (1993): Primitive dinosaur skeleton from Argentina and the early evolution of the Dinosauria. Nature. 361 (6407): 64–66.
Sereno, P. C., Martínez, Ricardo N., Alcober, Oscar A. (2013): Osteology of Eoraptor lunensis (Dinosauria, Sauropodomorpha). Basal sauropodomorphs and the vertebrate fossil record of the Ischigualasto Formation (Late Triassic: Carnian-Norian) of Argentina. Journal of Vertebrate Paleontology Memoir. 12: 83–179.
Sereno, P.C., Novas, F.E. (1992): The complete skull and skeleton of an early dinosaur. Science. 258 (5085): 1137–1140.
Sereno, P.C., Novas, F.E., Arcucci, A.B., Yu, C. (1988): New evidence on dinosaur and mammal origins from the Ischigualasto Formation (Upper Triassic, Argentina). Journal of Vertebrate Paleontology. 8 (3, supplement): 26A
Sterling J. Nesbitt, P. M. Barrett, Werning, S., et al (2012): The Oldest Dinosaur? A Middle Triassic Dinosauriform from Tanzania. Biology Letters. 9(1)
Stevens, K. A. (2013): The Articulation of Sauropod Necks: Methodology and Mythology. PLoS ONE 8(10): e78572.
Tang, F. et al. (2001): Biostratigraphy and palaeoenvironment of the dinosaur-bearing sediments in Lower Cretaceous of Mazongshan area, Gansu Province, China. Cretaceous Research. 22 (1): 115–129.
Taylor, M. P., Wedel, M. J. (2013a): The Effect of Intervertebral Cartilage on Neutral Posture and Range of Motion in the Necks of Sauropod Dinosaurs. PLoS ONE 8(10): e78214.
Taylor, M. P., Wedel, M. J. (2013b): Why sauropods had long necks, and why giraffes have short necks. PeerJ. 1: e36.
Thompson, R. S., Parish, J. C., Maidment, S. C. R., Barrett, P. M. (2011): Phylogeny of the ankylosaurian dinosaurs (Ornithischia: Thyreophora). Journal of Systematic Palaeontology 10(2): 301–312
Vinther, J., Nicholls, R., Kelly, Diane A. (2021): A cloacal opening in a non-avian dinosaur. Current Biology. 31 (4): R182–R183.
Vinther, J., Nicholls, R., Lautenschlager, S., Pittman, M., Kaye, T. G., Rayfield, E., Mayr, G., Cuthill, I. C. (2016): 3D Camouflage in an Ornithischian Dinosaur. Current Biology. 26 (18): 2456–2462.
Wedel, M. J., Taylor, M. P. (2013): Caudal Pneumaticity and Pneumatic Hiatuses in the Sauropod Dinosaurs Giraffatitan and Apatosaurus. PLoS ONE 8(10): e78213.
Weishampel, D. B., P. Dodson, and H. Osmolska, eds. (1990): The Dinosauria. 1st ed. Berkeley: University of California Press.
Xu, X., Forster, C. A., Clark, J. M., Mo, J. (2006): A basal ceratopsian with transitional features from the Late Jurassic of northwestern ChinaProc. R. Soc. B.2732135–2140
Yates, A. (2004): Anchisaurus polyzelus (Hitchcock): The smallest known sauropod dinosaur and the evolution of gigantism among sauropodomorph dinosaurs. Postilla. 230: 1–57.
Zanno, L. E. (2010a): Osteology of Falcarius utahensis (Dinosauria: Theropoda): characterizing the anatomy of basal therizinosaurs. Zoological Journal of the Linnean Society. 158 (1): 196–230.
Zanno, L. E. (2010b): A taxonomic and phylogenetic re-evaluation of Therizinosauria (Dinosauria: Maniraptora). Journal of Systematic Palaeontology. 8 (4): 503–543.
Zanno, L. E., Erickson, G. M. (2006): Ontogeny and life history of Falcarius utahensis, a primitive therizinosauroid from the Early Cretaceous of Utah. Journal of Vertebrate Paleontology. 26 (supp. 3): 143A.
