

Einführung
Text als pdf
Wohl kaum eine Gruppe ausgestorbener Tiere fasziniert Groß und Klein mehr als die Dinosaurier. Das Mesozoikum wird oft als das „Zeitalter der Dinosaurier“ bezeichnet, weil sich diese erstaunlichen Tiere in der späten Trias entwickelten und die Landschaft während der restlichen Zeit des Jura und der Kreidezeit über 140 Millionen Jahre beherrschten. Doch auch heute noch tummeln sich auf unserem Planeten eine Unzahl verschiedener Dinosaurierarten. Damit sind aber nicht Schildkröten, Eidechsen, Schlangen oder Krokodile gemeint – sondern die Vögel. Vom kleinsten Kolibri bis zum Vogelstrauß sind alle etwa 11.000 beschrieben heute lebenden Vögel waschechte Dinosaurier (Chiappe 2009, Currie 2023). Aber warum sind Vögel echte Dinosaurier, aber Echsen und Krokodile sind keine? Warum sollen Gattungen wie Effigia, die unserer Vorstellung eines Dinosauriers erstaunlich ähnlichsehen (Abb. 1), keine Dinosaurier, sondern eher mit Krokodilen verwandt sein (Nesbitt et al. 2006, Nesbitt 2007, Nesbitt 2011)? Und warum stehen Gattungen wie Silesaurus (Abb. 1), die für den Laien der Gattung Effigia so ähnlichsehen, den Dinosauriern näher, sind aber immer noch keine echten Dinosaurier (Nesbitt et al. 2010, Nesbitt 2011, Dzik 2003, Schmitt 2023)?

Abb.1: Effigia, Silesaurus und Grüner Leguan sind keine Dinosaurier. Kolibris aber schon. Doch warum?
Text als pdf
In den folgenden Beiträgen wollen wir erklären warum Vögel Dinosaurier sind und wollen dies anhand des Fossilberichts, der Kladistik und der Anatomie dieser Tiergruppen ergründen. Doch bevor wir dies tun, müssen wir zuerst einordnen was ein Dinosaurier überhaupt ist, denn das ist der erste Schritt, um zu erkennen, warum Kolibris Dinosaurier sind, Warane, Krokodile, Silesaurus und Effigia aber nicht.
Reptilien?
Dinosaurier gelten gemeinhin als Reptilien, während die Vögel eine eigene Klasse bilden, die auf den ersten Blick recht wenig mit der Klasse der Reptilien gemeinsam hat (Linneaus 1758, Abb. 2). Betrachtet man heute lebende Reptilien wie Schlangen und Schildkröten, so haben sie Merkmale, die sie klar von den Vögeln unterscheiden. Reptilien haben Schuppen, sind in der Regel wechselwarm, haben Zähne und ihre Gliedmaßen, sofern sie welche haben, stehen seitlich am Körper. Vögel hingegen haben Federn, einen zahnlosen Schnabel, können ihre Temperatur konstant halten und ihre Beine stehen senkrecht unterm Körper, während die Arme in Flügel umgewandelt sind. Betrachtet man also nur heute lebende Reptilien und Vögel, scheint eine Einteilung in zwei Klassen gerechtfertigt (vgl. Modesto & Anderson 2004, Gauthier 1994).

Abb. 2: betrachtet man die fünf traditionellen Wirbeltierklassen, so haben scheinbar Vögel und Reptilien völlig verschiedene Merkmale, die kaum auf eine nähere Verwandtschaft schließen lassen.
Gleichzeitig haben aber beide Klassen doch gewisse Gemeinsamkeiten: Auch wenn Vögel Federn haben, so haben sie an den Beinen reptilienartige Schuppen und Zehen mit Krallen. Außerdem legen beide hartschalige amniotische Eier. Die meisten Reptilien und auch die Vögel haben einen sog. diapsiden Schädelbau, d. h. sie haben, neben den Augen- und Nasenhöhlen zwei charakteristische Schläfenfenster auf jeder Schädelseite (Abb. 3). Ein oberes Schläfenfenster (Fenestra supratemporalis, Supratemporalfenster), das meist oben auf dem Schädeldach sitzt und ein unteres Schläfenfenster (Fenestra infratemporalis, Infratemporalfenster) in der Schädelseitenwand. Im Zuge der Evolution der Reptilien und Vögel wurde dieser diapside Schädelgrundtyp stark abgewandelt, lässt sich aber z. B. noch in der Embryonalentwicklung nachvollziehen. Weiterhin fiel Forschern auf, dass innerhalb der Reptilien Krokodile einige Gemeinsamkeiten mehr mit Vögeln haben – dies zeigt sich z. B. am Bau des Herzens, Details in der Schädelanatomie oder genetischer Merkmale (Benton 2007, Fastovsky & Weishampel 2021, Holtz 2007, Paul 2016, Prothero 2017, 2021, 2022, Prothero & Dott 2004, Brusatte 2019, Schmitt 2023).
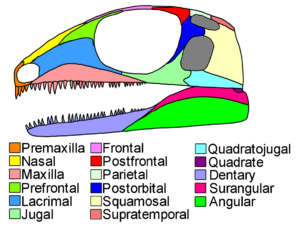
Abb. 3: Grundbau eines diapsiden Schädels. Man beachte die zwei hinteren Schläfenfenster, die grau markiert sind.
Wenn wir in dieses Bild nun ausgestorbene Vertreter wie die Dinosaurier zu Rate ziehen, verschwimmen die Grenzen zwischen Dinosauriern, die gewöhnlich zu den Reptilien gezählt werden und den Vögeln. In der Biologie wird heute darauf geachtet, dass bei der systematischen Einteilung der Lebewesen immer alle Nachfahren eines gemeinsamen Vorfahren zusammengefasst werden. Der Wissenschaftler nennt das „Monophylie“ (Abb. 4). Eine solche Einteilung in monophyletische Gruppen basiert auf evolutionären Neuheiten, also Apomorphien, die nur in Vertretern dieser monophyletischen Gruppe vorkommen. Solche monophyletischen Gruppen bezeichnen wir als Klade. Entsprechend wird diese Art der Einteilung in Lebewesen als Kladistik oder phylogenetische Systematik. Dabei ist jede untere Einheit Teil der nächst höheren. Die Kladistik ordnet Organismen in hierarchisch ineinander geschachtelte Gruppen ein, die ausschließlich durch evolutive Neuheiten (also Apomorphien) definiert sind. Mit größtmöglicher Objektivität entstehen so Systeme natürlicher Klassen, die sich in Verwandtschaftsbäume (Kladogramme) und diese wiederum in Stammbäume übersetzen lassen.
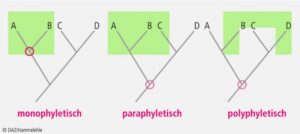
Abb. 4: Verwandtschaftsverhältnisse: mono-, poly- und paraphyletische Gruppen. Der rote Kreis bezeichnet die nächstliegende Verzweigung im Stammbaum, die allen Angehörigen der Gruppe (grün) gemeinsam ist. Paraphyletische und polyphyletische Gruppen schließen nicht alle Individuen eines Monophylums ein. Die moderne Taxonomie erlaubt nur monophyletische Gruppen.
Das Wort „Reptil“ erfüllt das Kriterium einer monophyletischen Abstammungsgemeinschaft jedoch nicht, weil es die Dinosaurier zwar ein-, aber die Vögel als ihre direkten Nachfahren ausschließt, und ist damit ein „paraphyletischer Begriff“, sprich: der Begriff ist ungenau und deshalb streng genommen falsch. Daher taucht in der Fachliteratur ein anderer Begriff auf, der sowohl alle Reptilien wie auch die Vögel einschließt: Sauropsida (Gauthier 1994, Abb. 5). Kurzum: Dinosaurier hatten zwar unter den Reptilien Vorfahren und auch nahe Verwandte wie zum Beispiel die Krokodile, doch waren sogar die ersten und urtümlichsten Dinosaurier den Vögeln schon viel ähnlicher als den klassischen Reptilien.
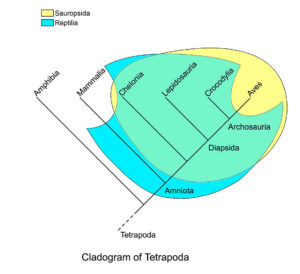
Abb. 5: Kladogramm der Tetrapoda, wobei Sauropsida und die Klasse Reptilia markiert sind. Die beiden überschneiden sich weitgehend, aber die traditionelle Klasse Reptilia umfasst die frühesten Amnioten sowie die säugetierähnlichen Reptilien, jedoch nicht die Vögel. Sauropsida umfasst die Vögel und die rezenten Reptiliengruppen (Schildkröten, Lepidosaurier [=Echsen und Schlangen] und Krokodile, sowie deren gemeinsamen Vorfahren), nicht jedoch die Synapsiden (= „säugetierähnliche Reptilien“), die zu den Vorfahren der modernen Säugetiere gehören. Nach dieser Definition bilden die Sauropsida eine monophyletische Klade.
Archosauria
Die Sauropsida teilt man in mehrere Kladen ein, deren zwei größten die Archosauromorpha und die Lepidosauromorpha sind (Ezcurra 2016, Martinelli et al. 2017, Butler et al. 2014, Gauthier et al. 1988, Nesbitt 2011; Abb. 6). Letztere vereinen die meisten klassischen Reptilien: allen voran Schlangen und Echsen, aber auch ausgestorbene Gruppen, z. B. die Plesiosaurier. Die Archosauromorpha lassen sich weiter unterteilen, so in die Archisauriformes, in der wiederrum die Archosauria eingeschachtelt sind (Gauthier 1994, Senter 2005, Fastovsky & Weishampel 2021, Nesbitt 2011, Ezcurra 2016). Zu den Archosauria gehören dann auch die Krokodile, Vögel, Flugsaurier und Dinosaurier, wie ihre Verwandten. Diese detaillierte Gabelung der verschiedenen Kladen wirkt für den Laien verwirrend, wird aber durch jeweilige Apomorphien definiert, die die entsprechenden Kladen aufweisen. Diese abgestufte Hierarchie von Apomorphien ist eine der besten Belege für den evolutionären Wandel, wo evolutionäre Neuheiten neue Gruppen definieren. Gerade bei einigen ursprünglichen Vertretern innerhalb einiger Kladen ist die Unterscheidung zu welcher Klade sie genau gehören nicht immer einfach. Wir werden leider nicht umhinkommen uns an diesem phylogenetischen Stammbaum zu orientieren, um endlich zu den Dinosauriern zu gelangen.
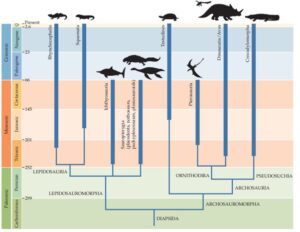
Abb. 6: Stammbaum der Sauropsida (bzw. hier als Diapsida), welche die zwei hauptäste der Lepidosauromorpha und Archosauromorpha vereint.
Welche Merkmale definiert die Klade der Archosaurier? Ein gemeinsames Merkmal aller Archosaurier liegt u. a. im Schädelbau (Abb. 7). Ausgehend vom diapsiden Schädeltyp mit den zwei Schläfenfenstern, findet sich vor der Augenhöhle das Antorbitalfenster. Bei modernen Vögeln und Krokodilen kam es hier jedoch zu starken Abwandlungen mit starker Spezialisierung. Die Krokodile haben zwar deutlich ausgeprägte Schläfenfenster, jedoch ist bei ihnen das Antorbitalfenster verschlossen (Abb. 8). Bei den Vögeln ist das Antorbitalfenster nur undeutlich, z. T. gar nicht von der Augenhöhle abgegrenzt und die Schläfenfenster sind klein und ebenfalls mit der Augenhöhle verschmolzen (Abb. 9). Das Antorbitalfenster reduzierte das Gewicht des Schädels, was u. a. auch ein Größenwachstum ermöglicht. Der Unterkiefer besitzt ursprünglich ebenfalls ein Fenster, welches als Mandibularfenster bezeichnet wird und bei den Krokodilen noch vorhanden ist (Abb. 7 und 8). Die Befestigung der Zähne in den Kieferknochen ist thecodont, d. h., die Zähne sitzen jeweils in einem eigenen Zahnfach (Alveole), wo sie durch Bindegewebe mit dem Knochen verbunden sind, und besitzen keine mehrteiligen Zahnwurzeln (Abb. 7). Dieses Merkmal ist bei Vögeln durch die Ausbildung des zahnlosen Schnabels verloren gegangen, findet sich aber z. b. bei einigen ausgestorbenen Vogelarten wie Ichthyornis.
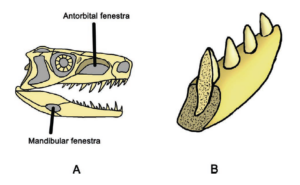
Abb. 7: Einige Synapomorphien der Archosauria. A. Sie haben zwei zusätzliche Öffnungen im Schädel: Das Antorbitalfenster und das Mandibularfenster. B. Die Befestigung der Zähne in den Kieferknochen ist thecodont, d. h., die Zähne sitzen jeweils in einem eigenen Zahnfach (Alveole), wo sie durch Bindegewebe mit dem Knochen verbunden sind, und besitzen keine mehrteiligen Zahnwurzeln. Im Gegensatz sitzen die Zähne der Lepidosaurier in einer Furche am inneren Rand des Kiefers.

Abb. 8: Schädel eines Krokodils. Sichtbar sind die beiden Schläfenfenster, während das Antorbitalfenster sekundär zurückgebildet ist. Erkennbar ist das Mandibularfenster im Unterkiefer.
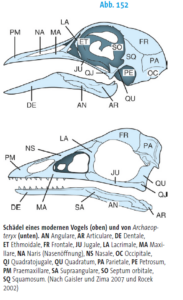
Abb. 9: Schädel eines modernen Vogels (oben) im Vergleich zu Archaeopteryx. Auffallend ist die Verschmelzung des Antorbitalfensters mit der Augenhöhle (getrennt werden beide Öffnungen durch das Lacrimale (LA).
Archosaurier haben zudem einen vierten Trochanter (Rollhügel) am Oberschenkelknochen. Der vierte Trochanter ist ein Höcker auf der posterior-medialen Seite der Mitte des Oberschenkelknochenschafts, das als Muskelansatz dient, hauptsächlich für den Musculus caudofemoralis longus, den Hauptretraktorschwanzmuskel, der den Oberschenkelknochen nach hinten zieht, und eine große Ansatzstelle für Muskeln am Oberschenkelknochen bietet (Nesbitt 2011, Fastovsky & Weishampel 2021, Holtz 2007, Paul 2016, Schmitt 2023; Abb. 10). Anders als bei den Lepidosauriern, wie den Schlangen, fehlt bei den Archosauriern das Jacobson-Organ, ein bei vielen Wirbeltieren ausgebildetes Geruchsorgan (Poncelet & Shimeld 2020, Abb. 11). Diese Apomorphien der Archosaurier finden sich natürlich auch bei den Dinosauriern.

Abb. 10: Links: Das linke Bein eines großen Entenschnabelsauriers mit einer eingezeichneten roten Linie, die den ungefähren Verlauf des Musculus caudofemoralis longus zeigt, der am vierten Trochanter endet. Rechts: Rekonstruktion des Musculus caudofemoralis longus bei Tyrannosaurus rex.
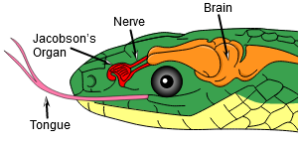
Abb. 11: Jacobson-Organ bei der Schlange. Dieses Merkmal fehlt den Archosauriern.
Avemetatarsalia und Ornithodira
An der Basis der Archosauriforma stehen Gattungen wie Euparkeria oder Proterosuchus (Gauthier 1986, Sookias & Butler 2013, Nesbitt 2011, Ezcurra & Butler 2015, Welman 1998), die schon einige, aber nicht alle der Apomorphien der Archosaurier teilen (Abb. 12).
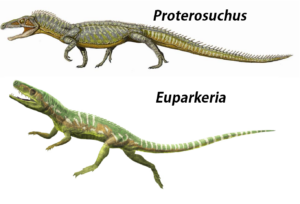
Abb. 12: Proterosuchus und Euparkeria, die an der Basis der Archosauriforma stehen
Die Archosaurier selbst lassen sich in zwei Kladen einteilen: Die Crurotarsi, bzw. Pseudosuchia, welche die Krokodile und ihre vielfältigen ausgestorbenen Verwandten einschließen und die Avemetatarsalia, welche u. a. die Dinosauromorpha und Pterosauromorpha, aber auch einige ursprünglichere Kladen einschließen (Fastovsky & Weishampel 2021, Holtz 2007, Paul 2016, Prothero 2017, 2021, 2022, Prothero & Dott 2004, Nesbitt & Norell 2007, Nesbitt 2007, 2011, Nesbitt et al. 2017, Brusatte 2019, Brusatte et al. 2010a,b, Sereno 1991, Gauthier 1986, Gauthier & de Queiroz 2001, Cau 2018, Abb. 13).
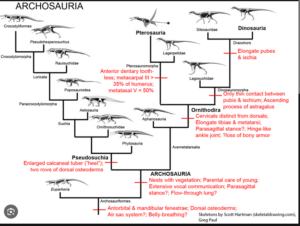
Abb. 13: Kladogramm der Archosaurier mit einigen Schlüsselmerkmalen
Zu den ursprünglichen Avemetatarsalia gehören u.a. Aphanosaurier, die an der Basis der Avemetatarsalia stehen (Nesbitt 2011, Nesbitt et al. 2017). Die Aphanosaurier wiesen Merkmale sowohl der Avemetatarsalia, aber auch der Crurotarsi auf, was darauf hindeutet, dass sie die älteste und ursprünglichste bekannte Gattung der Avemetatarsalier sind, zumindest was ihre Position im Stammbaum der Archosaurier betrifft. Die Dinosauromorpha und Pterosauromorpha vereinigt man als Ornithodira (Abb. 13).
Ein Unterscheidungsmerkmal zwischen Crurotarsi und Avemetatarsalia ist der Bau des Fußgelenks, genauer des Sprunggelenks (Fastovsky & Weishampel 2021, Prothero 2017, 2021, 2022, Abb. 14). Das Sprunggelenk ist das Verbindungsgelenk zwischen dem Unterschenkel und dem Fuß. Am Sprunggelenk sind die oberen Fußwurzelknochen Astragalus (Sprungbein) und Calcaneus (Fersenbein) beteiligt. Die Avemetatarsalia zeichnen sich durch einen großen Astragalus und kleinen Calcaneus aus. Der Astragalus und der Calcaneus sind beide mit dem Ende des Schienbeins und Wadenbein verschmolzen. Das Scharnier des Gelenks befindet sich so zwischen Astragalus und Calcaneus und der zweiten Reihe der Fußwurzelknochen. Man spricht hierbei von einem Mesotarsalgelenk, also eine Gelenkung in der „Mitte“ der Fußwurzel und findet sich bei allen Flugsauriern, Dinosauriern und damit Vögeln. Diese Bauweise ermöglichte nicht nur über eine bessere Beweglichkeit, sondern erlaubt den Tieren eine Fortbewegung auf ihren Zehen. Alle Dinosaurier laufen auf den Finger- und Zehenspitzen und nicht auf den Handflächen und Fußsohlen. Das gilt selbst für die schwersten Riesensauropoden, die immer noch auf den Zehenspitzen gehen, deren Zehenknochen aber aufgrund ihres Gewichts zu stummeligen Säulen zusammengequetscht sind.
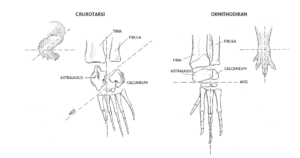
Abb. 14: Knöchel der Crurotarsi und Ornithodira (bzw. Avemetatarsalia) im Vergleich. Erläuterungen siehe Text.
Lange Zeit waren die Crurotarsi die bevorzugte Bezeichnung für den Krokodilzweig, da sie sich durch eine ausgeprägte Knöchelkonfiguration auszeichneten, bei der das Scharnier des Winkels zwischen dem Calcaneus und dem Astragalus verläuft, wobei der Calcaneus größer ist als bei den Avemetatarsalia. Der Astragalus ist nur mit Ende des Schienbeins verwachsen und das Gelenk verläuft zwischen Calcaneus und Wadenbein. Diese Anordnung führt dazu, dass Krokodile und ihre Verwandten mit der gesamten Fußsohle auftreten. Neuere Analysen haben jedoch ergeben, dass einige Gruppen (die Phytosaurier) primitiver sind als die übrigen Mitglieder des Krokodilzweigs, so dass der Krokodilzweig jetzt als Pseudosuchia bezeichnet wird und die Phytosaurier entweder direkt außerhalb der Pseudosuchia oder in anderen Fällen als primitive Archosauromorphe direkt außerhalb der Archosauria eingeordnet werden (Nesbitt 2011).
Dinosaurier
Innerhalb der Ornithodira gehören die Dinosaurier, nebst einigen weiteren Taxa zu den Dinosauromorpha (Benton 1985, Sereno 1991, Nesbitt 2011, Cau 2018, Gauthier 1986, Ezcurra et al. 2020). Ihre Schwestergruppe sind die Pterosauromorpha. Die Dinosauromorpha haben zwar einige, jedoch nicht alle Apomorphien der Dinosaurier. Für den Nicht-Spezialisten sehen sie oberflächlich wie Dinosaurier aus, sind es aber noch nicht ganz. Einige kleinere Gattungen wie Lagosuchus, Marasuchus und Silesaurus (Abb. 1 und 15) gehören dazu (Romer 1971, Agnolin & Ezcurra 2019, Sereno & Arcucci 1994, Nesbitt et al. 2010, Nesbitt 2011, Dzik 2003, Schmitt 2023). Aus solchen oder ähnlichen Formen gingen dann die Dinosaurier mit ihren eigenen Apomorphien hervor. Also auch hier, wie bei den Archosauromorpha und Archosauriern: Jeder Dinosaurier gehört zu den Dinosauromorpha, aber nicht jeder Dinosauromorpha ist ein Dinosaurier. Welche Apomorphien kennzeichnen die Dinosaurier?
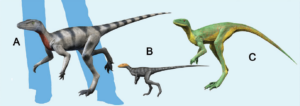
Abb. 15: einige Dinosauromorpha: (A) Saltopus, (B) Marasuchus, (C) Lagerpeton.
Als die allerersten bekannten Dinosaurier nur Megalosaurus (Buckland 1825) und Iguanodon (Mantell 1825) und einige andere waren, benannte und diagnostizierte Richard Owen sie als eine Gruppe riesiger ausgestorbener Reptilien mit einer Reihe charakteristischen Merkmalen (Owen 1842). Als die Zahl neuer Dinosaurierfunde in den späten 1800er und frühen 1900er Jahren rapide zunahm, wurde diese Definition geändert. Als Samuel Wendell Williston 1925 Osteology of the Reptiles veröffentlichte (Williston 1925), waren die Dinosaurier klar von anderen Reptiliengruppen wie den Meeresreptilien, den Flugsauriern und anderen abgegrenzt.
Im Jahr 1878 erkannte Othniel Charles Marsh vier Gruppen von Dinosauriern: Sauropoden, Theropoden, Ornithopoden und Stegosaurier, Gruppen, die auch heute noch gültig sind. Aber nur wenige dieser Autoren gaben eine genaue anatomische Diagnose dessen, was einen Dinosaurier ausmacht (Prothero 2022).
Im Jahr 1888 erkannte der britische Paläontologe Harry Govier Seeley (Seeley 1888) zwei Gruppen von Dinosauriern an: die „echsenartigen“ Dinosaurier oder Saurischia (mit denen er die Theropoden und Sauropoden zusammenfasste) und die „vogelartigen“ Dinosaurier oder Ornithischia (zu denen die meisten pflanzenfressenden Dinosaurier außer den Sauropoden gehören). Diese Vorstellungen setzten sich in den nächsten 130 Jahren durch, so dass sich die meisten Paläontologen darauf einigten, dass ein Fossil zu einer dieser beiden Gruppen gehören musste, um ein Dinosaurier zu sein (Abb. 16).
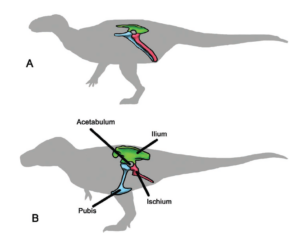
Abb. 16: die beiden Hauptgruppen der Dinosaurier: A. Vogelbeckendinosaurier („Ornithischia“); B. Echsenbeckendinosaurier (Saurischia)
Robert Bakker und Peter Galton zeigten 1974 (Bakker & Galton 1974) jedoch, dass die Dinosaurier eine Reihe einzigartiger anatomischer Merkmale aufweisen, die belegen, dass sie eine einzige natürliche Gruppe sind und sich nicht unabhängig voneinander aus verschiedenen frühen Archosauriern, den Thecodontia, entwickelt haben.
Als mehr und mehr Fossilien gefunden wurden, schienen die Unterschiede zwischen den beiden Gruppen der Dinosaurier, basierend auf ihrer Hüftstruktur konsistent zu sein, und die einzelnen Gruppen (Sauropoda, Theropoda usw.) funktionierten weiterhin gut. Die Saurischia waren die Dinosaurier mit dem Echsenbecken, bei denen das Schambein in der Hüftregion nach vorne zeigte. Bei den Ornithischia, den „Vogelbeckendinosauriern“, war zumindest ein Teil des Schambeins nach hinten verlagert, parallel zum hinteren Knochen der Hüftregion, dem Sitzbein. Hier sei schon mal die Ironie erwähnt, dass die Vögel sich aus den Theropoden (Raubsaurier) ableiten, die ein Echsenbecken hatten. Die Einteilung in Echsenbecken- und Vogelbeckendinosaurier, die ja schon seit 1888 existiert, war weit vor der Zeit, als die Verwandtschaft zwischen Dinosauriern und Vögeln eindeutig war. Die Ähnlichkeit des Beckens der Vogelbeckendinosaurier zu den Vögeln ist auch nur oberflächlich und kein Indiz für ein Abstammungsverhältnis. Dazu mehr in den nächsten Beiträgen.
Die von Seeley diagnostizierten Hauptunterordnungen der Dinosaurier beantworteten jedoch nicht die Frage, wie die Gruppen innerhalb der Saurischia und Ornithischia miteinander verbunden sind. Noch in den frühen 1970er Jahren waren Paläontologen nicht sicher, ob Saurischia und Ornithischia zu den Dinosauria zusammengefasst werden konnten. In seinem 1955 erschienenen Lehrbuch „Evolution of the Vertebrates“ schrieb Edwin Colbert, dass „der Begriff zwei verschiedene Reptilienordnungen umfasst. Folglich ist das Wort Dinosaurier jetzt ein praktischer umgangssprachlicher Name, aber kein systematischer“ (Colbert 1955).
Dieses traurige Missverständnis mag typisch für die Denkweise in den Jahren 1950er und 1960er gewesen sein, als die ersten beiden Auflagen von Colberts Buch veröffentlicht wurden (Colbert 1955, 1961), aber leider blieb der Text auch in der letzten Auflage von 2001 unverändert, als diese Idee durchschlagend entkräftet wurde (Colbert et al. 2001). Das lag daran, dass Biologen und Paläontologen in den 1970er und 1980er Jahren begannen, Methoden der Kladistik als Klassifizierungsmethode anzuwenden, die nach einzigartigen evolutionären Neuerungen suchte, die natürliche Gruppen definierten, und sich von sog. „wasteback“-Gruppen entfernte, die unnatürliche Zusammenstellungen von nicht verwandten Tieren waren.
Mittlerweile sind eine Reihe anatomischer Merkmale bekannt, die Dinosaurier definieren (vgl. Fastovsky & Weishampel 2021, Holtz 2007, Paul 2016, Prothero 2017, 2021, 2022, Prothero & Dott 2004, Bakker & Galton 1974, Brusatte et al. 2010b, Gauthier 1986, Langer et al. 2010, Nesbitt et al. 2017, Sereno 1997, 1999, Sereno et al. 1993, Brusatte 2019, Schmitt 2023, Schweitzer et al. 2021 Abb. 17).
Eine der bedeutendsten wurde 1985 von Jacques Gauthier definiert (Gauthier 1986). Dieses Betrifft die Hüftgelenkpfanne, also jenem Bereich des Beckens, in dem der Oberschenkelknochen steckt. Bei vielen Landtieren ist die Hüftgelenkpfanne verschlossen, bei fast allen Dinosauriern findet sich jedoch eine Öffnung (das Acetabulum), die direkt durch die Hüftknochen geht.
Schließlich gibt es nur drei Wirbel, die mit dem oberen Teil der Hüftknochen verschmolzen sind, die Wirbelsäule mit den Hinterbeinen verbinden und ein Kreuzbein bilden. Andere Tiere haben weniger oder mehr Wirbel in ihren Hüften.
Dinosaurier haben Beine, die unter dem Körper stehen (während sie sich bei anderen Reptilien seitlich am Körper befinden). Diese Stellung der Beine erlaubt nicht nur eine effizientere Fortbewegung, sondern ermöglichte auch den Gigantismus vieler Dinosauriergruppen.
Alle Archosaurier besitzen, wie erwähnt, einen vierten Trochanter, aber bei Nicht-Dinosaurier-Archosauriern wie Krokodilen ist dieser Fortsatz abgerundet und symmetrisch. Bei den Dinosauriern ist der Trochanter deutlich asymmetrisch. Es wird vermutet, dass diese Asymmetrie auch eine Rolle bei bei der Unterstützung ihrer aufrechten Haltung gespielt haben könnte.
Am Oberarmknochen (Humerus) weisen Dinosaurier einen länglicher deltopectoralen Kamm auf: An diesem setzen zwei Muskeln ans, der Deltamuskel (bei uns der Muskel an der Außenseite der Schulter) und der Pectoralis (die Brustmuskeln), daher der Begriff „deltopectoral“. Eine längere Ansatzstelle für diese Muskeln deutet auf eine größere Kraft in den Vordergliedmaßen hin.
Auch an ihren Schädeln kann man einige spezielle Alleinstellungsmerkmale festmachen, die die Dinosaurier von anderen Reptilien unterscheiden: das Postfrontale, ein kleiner Deckenknochen des Schädeldachs, fehlt ihnen allen, im Gaumen überlappt das Ectopterygoid das Flügelbein, der Kopf des Kiefergelenks (Os quadratum) ist in seitlicher Ansicht exponiert, und die Posttemporalöffnung am Hinterhaupt ist verkleinert.
Andere anatomische Eigenheiten der Dinosaurier waren eine rückwärtig orientierte Schultergelenkpfanne, asymmetrische Hände mit verkürzten vierten und fünften Fingern (bei vielen Arten fehlten diese auch ganz), der Cnemialkamm (eine kammartige Erhebung am oberen Ende des Schienbeins), ein aufwärts gerichteter Fortsatz am Astragalus sowie ein S-förmig gebogener mittlerer Mittelfußknochen. Viele dieser Merkmale, die zumindest die früheren Dinosaurier aus der Trias und dem frühen Jura auf jeden Fall noch teilen, gingen jedoch bei höher entwickelten Dinosauriern im Laufe der Zeit verloren oder haben sich in anderer Weise verändert. Der anatomische Bauplan eines Dinosauriers aus der Kreidezeit kann also bereits stark von dem eines triassischen Dinosauriers abweichen.
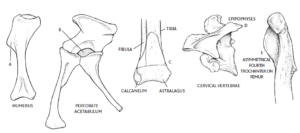
Abb. 17: einige Synapomorphien der Dinosaurier. a) verlängerter deltopectoraler Kamm am Humerus (erhöht die Kraft der Unterarme); b) perforierte Hüftpfanne; c) Fibula berührt 30 % des Astragalus (Teil des Mesotarsalknöchels); d) Epipophysen an den Halswirbeln (verringert die Rotation um den Hals); und e) asymmetrischer vierter Trochanter am Oberschenkelknochen.
Weitere Merkmale der Dinosaurier betreffen das Herz-Kreislaufsystem und den Bau der Lunge, diese werden aber in späteren Kapiteln, besonders mit der Verwandtschaft der Vögel, diskutiert.
Nun haben wir es endlich geschafft: Sauropsida, Archosauromorpha, Archisauriformes, Archosaurier, Avemetatarsalia, Ornithodira, Dinosauromorpha, Dinosaurier. Entschuldigt, wenn ich einige Kladen dazwischen vergessen oder übersprungen habe. Wer bei diesem verwirrenden Name-Game bis hierher mitgekommen ist, dem Gebührt Dank und Ehre. Wir wissen jetzt was ein Dinosaurier ist, bzw. an welchen Merkmalen man einen erkennt. Aber wirklich bei den Dinosauriern und erst recht bei den Vögeln sind wir noch nicht angekommen. Wenn wir über den Übergang von Dinosauriern zu Vögeln reden, dann ist nicht nur ihre Einordnung im Stammbaum interessant, sondern auch welche entscheidenden evolutionären Übergänge stattgefunden haben. Und eines sei versichert: diese sind sehr gut dokumentiert und belegt. Doch in der nächsten Episode soll es erstmal um die Nicht-Vogel-Dinosaurier gehen. Denn auch diese haben nicht nur eine erstaunliche Vielfalt, sondern auch einige interessante evolutionäre Übergänge. Wer also wissen möchte wie aus einigen fleischfressenden Dinosauriern reine Vegetarier wurden und wie aus kleinen Dinos so riesige Ungetüme wie Brachiosaurus und Argentinosaurus, der schaut sich die nächste Episode zu diesem Thema an.
Literatur
Agnolin, F. L., Ezcurra, M. D. (2019): The Validity of Lagosuchus Talampayensis Romer, 1971 (Archosauria, Dinosauriformes), from the Late Triassic of Argentina. Breviora. 565 (1): 1–21.
Bakker, R., Galton, P. (1974): Dinosaur Monophyly and a New Class of Vertebrates. Nature. 248. 10.1038/248168a0.
Benton, M. J. (2007): Paläontologie der Wirbeltiere. Übersetzung der 3. englischsprachigen Auflage. Verlag Dr. Friedrich Pfeil, München
Brusatte, S. L. (2019): The Rise and Fall of the Dinosaurs: A New History of a Lost World. William Morrow, NY
Brusatte, S. L., Benton, M. J., Desojo, J. B., Langer, M. C. (2010a): The higher-level phylogeny of Archosauria (Tetrapoda: Diapsida). Journal of Systematic Palaeontology. 8 (1): 3–47.
Brusatte, S. L., Nesbitt, S. J., Irmis, R. B., et al. (2010b): The origin and early radiation of dinosaurs. Earth-Science Reviews, 101, 68–100.
Buckland W. (1824): Notice on the Megalosaurus or great fossil lizard of Stonesfield. Transactions of the Geological Society of London, 1(2): 390–396.
Butler, R. J., Scheyer, T. M., Ezcurra, M. D. (2014): The Origin and Early Evolution of Sauria: Reassessing the Permian Saurian Fossil Record and the Timing of the Crocodile-Lizard Divergence. PLOS ONE. 9 (2): e89165.
Cau, A. (2018): The assembly of the avian body plan: a 160-million-year long process. Bollettino della Società Paleontologica Italiana. 57 (1): 1–25.
Chiappe, L. M. (2009): Downsized Dinosaurs: The Evolutionary Transition to Modern Birds. Evo Edu Outreach 2, 248–256
Colbert, E. H. (1955): Evolution of Vertebrates, 1st edition
Colbert, E. H. (1961): Evolution of Vertebrates, 2nd. edition
Colbert, E. H., Morales, M., Minkoff, E. C. (2001): Evolution of Vertebrates, 5th. Edition
Currie, P. J. (2023): Celebrating dinosaurs: their behaviour, evolution, growth, and physiology. Canadian Journal of Earth Sciences. 60(3): 263-293.
Dzik, J. (2003): A beaked herbivorous archosaur with dinosaur affinities from the early Late Triassic of Poland. Journal of Vertebrate Paleontology. 23 (3): 556–574.
Ezcurra, M. D. (2016): The phylogenetic relationships of basal archosauromorphs, with an emphasis on the systematics of proterosuchian archosauriforms. PeerJ. 4: e1778.
Ezcurra, M. D., Butler, R. J. (2015): Taxonomy of the proterosuchid archosauriforms (Diapsida: Archosauromorpha) from the earliest Triassic of South Africa, and implications for the early archosauriform radiation. Palaeontology. 58 (1): 141–170.
Ezcurra, M. D., Nesbitt, S. J., Bronzati, M., Dalla Vecchia, F. M., Agnolin, F. L., Benson, R. B. J., Brissón Egli, F., Cabreira, S. F., Evers, S. W., Gentil, A. R., Irmis, R. B. (2020): Enigmatic dinosaur precursors bridge the gap to the origin of Pterosauria. Nature. 588 (7838): 445–449.
Fastovsky, D. E., Weishampel, D. B. (2021): Dinosaurs: A Concise Natural History (4th ed.). Cambridge university Press, Cambridge.
Gauthier, J. A. (1986): Saurischian monophyly and the origin of birds. Memoirs of the California Academy of Sciences. 8: 1–55.
Gauthier, J. A. (1994): The diversification of the amniotes. In Prothero, D.R., Schoch, R.M. (eds.). Major Features of Vertebrate Evolution. pp. 129–159.
Gauthier, J. A., de Queiroz, K. (2001): Feathered dinosaurs, flying dinosaurs, crown dinosaurs, and the name „Aves“. pp. 7–41. In Gauthier, J. and L. F. Gall (eds.), New Perspectives on the Origin and Early Evolution of Birds: Proceedings of the International Symposium in Honor of John H. Ostrom. New Haven: Peabody Museum of Natural History, Yale University.
Gauthier, J. A., Kluge, A. G., Rowe, T. (1988): Amniote phylogeny and the importance of fossils. Cladistics. 4 (2): 105–209.
Holtz, T. R. (2007): Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages
Langer, M. C., Ezcurra, M. D., Bittencourt, J. S., Novas, F. E. (2010): The origin and early evolution of dinosaurs. Biological Reviews, 85, 55–110
Linnaeus, C. (1758): Systema naturae per regna tria naturae: Secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis [System of Nature through the Three Natural Kingdoms, According to classes, orders, genera, species, with characters, differences, synonyms, places] (in Latin)
Mantell G. A. (1825): Notice on the Iguanodon, a newly discovered fossil reptile, from the sandstone of Tilgate forest, in Sussex. Philosophical Transactions of the Royal Society, 115: 179–186.
Martinelli, A. G., Francischini, H., Dentzien-Dias, P. C., Soares, M. B., Schultz, C. L. (2017): The oldest archosauromorph from South America: postcranial remains from the Guadalupian (mid-Permian) Rio do Rasto Formation (Paraná Basin), southern Brazil. Historical Biology. 29 (1): 76–84.
Modesto, S. P., Anderson, J. S. (2004): The phylogenetic definition of Reptilia. Systematic Biology. 53 (5): 815–821.
Nesbitt, S. J. (2011): The early evolution of archosaurs: relationships and the origin of major clades. Bulletin of the American Museum of Natural History, 352, 1–292
Nesbitt, S. J. (2007): The anatomy of Effigia okeeffeae (Archosauria, Suchia), theropod-like convergence, and the distribution of related taxa. Bulletin of the American Museum of Natural History, 302:84
Nesbitt, S. J., Butler, R. J., Ezcurra, M. D., et al. (2017): The earliest bird-line archosaurs and the assembly of the dinosaur body plan. Nature, 544, 484–487.
Nesbitt, S. J, Norell, M. A. (2006): Extreme convergence in the body plans of an early suchian (Archosauria) and ornithomimid dinosaurs (Theropoda). Proceedings of the Royal Society B: Biological Sciences. 273 (1590): 1045–1048.
Nesbitt, S. J., Sidor, C. A., Irmis, R. B., Angielczyk, K. D., Smith, R. M. H., Tsuji, L. M. A. (2010): Ecologically distinct dinosaurian sister group shows early diversification of Ornithodira. Nature. 464 (7285): 95–98.
Owen R. (1842): Report on British fossil reptiles, part II. Report of the British Association for the Advancement of Science, 11:32–37.
Paul, G. S. (2016). The Princeton Fieldguide to Dinosaurs. 2nd Edition. Princeton University Press
Poncelet, G., Shimeld, S. M. (2020): The evolutionary origin of the vertebrate olfactory system. Open Biol. 10:200330.
Prothero, D. (2017): Evolution – What the Fossils say and why it matters. Second edition. New York: Columbia University Press
Prothero, D. (2021): The Evolving Earth. Oxford University Press
Prothero, D. (2022): Vertebrate Evolution. From Origins to Dinosaurs and beyond. CRC Press, Kapitel 11, 12, 14
Prothero, D. & Dott, (2004): Evolution of the Earth, Seventh edition. McGrawHill
Romer, A. S. (1971): The Chañares (Argentina) Triassic reptile fauna. X. Two new but incompletely known long-limbed pseudosuchians. Breviora. 378: 1–10.
Schmitt, A. (2023): Großartige Giganten. Den letzten Geheimnissen der Dinosaurier auf der Spur. Dtv Verlagsgesellschaft
Schweitzer, M. H., Schroeter, E. R., Czajka, C. D. (2021): Dinosaurs How We KNow What We Know. CRC Press
Seeley, H. G. (1888): On the classification of the Fossil Animals commonly named Dinosauria. Proceedings of the Royal Society of London. 43 (258–265): 165–171.
Senter, P. (2005): Phylogenetic taxonomy and the names of the major archosaurian (Reptilia) clades. PaleoBios. 25 (2): 1–7.
Sereno, P. C. (1991): Basal archosaurs: phylogenetic relationships and functional implications. Journal of Vertebrate Paleontology Memoir 2, 11(4, Supplement):1–53.
Sereno, P. C. (1997): The origin and evolution of dinosaurs. Annual Reviews of Earth and Planetary Sciences. 25: 435–490.
Sereno, P. C. (1999): The evolution of dinosaurs. Science. 284 (5423): 2137–2147.
Sereno, P. C., Arcucci, A. B. (1994): Dinosaurian precursors from the Middle Triassic of Argentina: Marasuchus lilloensis, gen. nov. Journal of Vertebrate Paleontology. 14 (1): 53–73.
Sereno, P.C.; Forster, C.A.; Rogers, R.R.; Monetta, A.M. (1993): Primitive dinosaur skeleton from argentina and the early evolution of Dinosauria. Nature Research. 361 (6407): 64–66.
Sookias, R. B., Butler, R. J. (2013): Euparkeriidae. Geological Society, London, Special Publications. 379 (1): 35–48.
Welman, J. (1998): The taxonomy of the South African proterosuchids (Reptilia, Archosauromorpha). Journal of Vertebrate Paleontology. 18 (2): 340–347.
Williston, S. W. (1925): Osteology of the Reptiles
