
Especially in parts two and three, we have explained the biological justification of why there are two sexes and why this makes sense. In nature, there are different mechanisms of how the sexes are determined (XX / XY, ZW / ZZ, XX / X0, environmental sex determination). In addition to separate sexes, there are so-called hermaphrodites, i.e. those that combine both sexes in one body (e.g. in many flowering plants or snails).
These forms of sexual determination correspond to the normal distribution of living organisms. Of course, there are deviations from the „norm“. Terms such as „normal“, „abnormal“, „untypical“ and „deviations“ are to be understood in biology as value-neutral terms that do not involve any discrimination. They only say that there are individuals who do not correspond to the majority. This is also true for sex distribution. It often comes either to chromosomal abnormalities or hormonal disorders (e.g., wasted gonads or genitalia). One speaks of intersexuals, a term introduced by the geneticist Richard Goldschmidt (1878-1958). Some people also bring the concept of the „third sex“ into the discussion. However, this is insufficient because no new type of germ cell is formed and many intersex people are sterile (KUTSCHERA 2018: 213-219). A study by BLACKLESS et al. (2000), who collected the results from 20 nations, was able to evaluate that one out of 2,000 newborn babies cannot be clearly assigned to a sex. This means that they are very rare. In addition, all intersex types are integrated here, of which there are quite a few forms. We want to introduce them gradually (according to KUTSCHERA 2018, BLUMBERG 2009, GILBERT 2006).
Real hermaphrodites
These people usually have two X chromosomes (XX) but have unknownly developed ovarian and testicular tissue in the gonads. The genitals show a continuum from typical woman to typical man. About the emergence of this intersex form is virtually nothing known, and the inheritance is unclear. To recognize „true hermaphroditism“, one must look at the gonadal tissue under the microscope.
Pseudohermaphrodites
In pseudohermaphrodites, only one gonadal tissue is developed, but the attached sex organs show atypical distributions.
For example, the phenomenon of oviduct persistence exists. In this case, no anti-Müllerian hormone is produced or the corresponding receptors on the cells to which the anti-Müller hormone can dock are missing. Although testicles develop, but they remain in the body cavity, so do not wander into the scrotum „outside“ (cryptorchidism). But this syndrome also develops outer genitalia and a vas deferens. But since the anti-Müllerian hormone is not present, or cannot work, parts of Müllerians’s duct remain and a small (dysfunctional) uterus is developed. The development of puberty is normal.
In androgen insensivity syndrome (AIS), testicles are formed and testosterone is produced. However, it lacks the receptors to which the testosterone can dock and thus becomes functionless. AIS can vary in severity; in its most pronounced form, the external genitalia become female. However, the vagina ends blindly, as no uterus is formed. The phenotype appears female, it also forms a breast tissue. Body hair is practically non-existent (see Fig. 1).
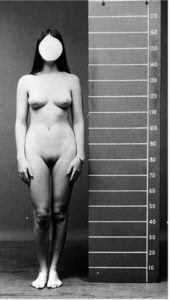
Fig.1: androgen insensivity syndrome (AIS). Source: GILBERT 2006
Another syndrome of XY pseudohermaphrodites is 5-alpha reductase deficiency syndrome, a form of androgen synthesis disorder that lacks the enzyme 5-alpha reductase. This enzyme can make dihydrotestosterone from testosterone, which is responsible for the development of male genitals. If the enzyme is missing, the genitals are not completely masculinized.
The Klinefelter syndrome is characterized by the fact that these persons have two X chromosomes and one Y chromosome (XXY). At birth, these people have no visible anomalies, but at puberty, they grow female breast tissue, the testes remain underdeveloped and they are sterile.
Another example is the Turner syndrome. These people lack the second X chromosome, so they are X0. The persons have a female body and are of small stature. Their ovaries, however, are greatly reduced or absent. The genital tract is that of a non-sexually mature female. They too are sterile.
We have already addressed the XX men in part three. In XX males, the SRY gene migrated to one of the X chromosomes, causing the individual to develop male genitals despite of two X chromosomes. The frequency is 1: 20,000. This makes it much less common than Klinefelter syndrome, but its appearance is similar (due to the presence of two X chromosomes). They too are sterile.
In congenital adrenal hyperlasia (CAH), people have 2 X chromosomes (XX) and develop ovaries. But their external genitalia become male, because during the embryonic development the hormone balance of the adrenal cortex is disturbed, which in turn has an influence on the formation of sex hormones.
Another form of intersexuality is the Swyer syndrome, in which the Y chromosome is mutated and thus forms a female phenotype (similar to the AIS discussed above, these individuals are sterile).
Other forms of pseudohermaphrodites include phenomena such as prolonged clitoris, micropenis, and chromosomal abnormalities such as XXX, XXXX, XXXY, and XXXXY. These should not be discussed here.
Intersexuals cannot be called the third sex. They are either often sterile or are not fully developed in the sense of sexual determination. For the definition of the sexes, however, the functional necessity of germ cell production and reproduction is a prerequisite. There are a number of mutations and chromosomal abnormalities in humans (and living things in general), regardless of gender: albinism, trisomy 21 („Down syndrome“), lactose intolerance, sickle cell anemia, etc. Such „anomalies“ are part of the process of genetic diversity. Why should sexes be excluded? People with trisomy 21 are also not forced into a new race of men, but are accepted as human beings (terms like „Mongoloid“ or „Mongo“, on the other hand, are pejorative, offensive and discriminatory and therefore cannot be used!).
At birth, gender neutral?
By arbitrary decision intersexual people were assigned in one of these two sexes by surgery and hormone treatment. But one has – rightly! – gotten away from this practice. Surgeries on intersex people are based on the thoughts of the sexologist and psychologist John Money, who argued that one is gender-neutral at birth and one can be educated to male or female gender. Money advised Bruce Reimer’s parents to undergo a sex change operation in 1967, after his circumcision had irreparably injured his penis. At the age of 22 months, the testicles were removed and labia were formed, from the age of 12, female sex hormones were administered. But Bruce, now called Brenda, did not accept the gender roles he had been taught. Bruce / Brenda wanted to reverse his sex change from the age of 14 and called himself from the time David. He committed suicide in 2004 (see KUTSCHERA 2018). KUTSCHERA 2018 draws a connection between the case John Money / Bruce Reimer and the gender ideology. I cannot judge this because it has little relevance for the other articles, but I cannot deny it or prove it is the case (Kutschera’s drawn connection between gender ideology and Marxism, however, are pure nonsense). The fact remains that science rejects John Money’s theories and these are refuted, especially in intersex. For us, the demands of intersex associations are decisive, how to deal with the issue of intersexuality. The Intersexual Association of North America (ISNA) writes that they are only for an operative measure if there is a) medically necessary (e.g. if it can save a person’s life) and b) the intersex person is informed of all risks and benefits. But they refuse an optional surgery (mostly on children) without the consent of the patient. Source
They also refuse to rais intersex children for any „third gender“ (which does not exist). The ISNA does not want to make any rules in which direction the children should be educated. They know that being an intersex person or having an intersex child is difficult. What they can do – and what one can do in general – is to provide proper information about intersex, psychological or medical advice and assistance, as well as to establish contacts with other intersex people. Source:
A demand that can and must be supported. Because that helps more than the invention of additional genders.
For example, the adaptation of a hypospadical penis (developmental defect of the urethra whose opening is further down the penis) is associated with years of surgery and pain that leave an otherwise functional penis in a scarred and dysfunctional state. A study of 20 heterosexual men who have micropenis and have not been surgically altered showed that 15 of them had successful sex with women. The partnerships were long-lasting and stable, the men self-confident and eager to experiment. One was even a father (WOODHOUSE 1998 quoted in BLUMBERG 2009).
Although men with cloacal or classic bladder exstrophy (incidence: 1 in 400,000 births) are not considered intersex in the classical sense, they have an abnormal pelvic region where men have either no or a very deformed penis. A standard method was to turn men with this disease into women. In a study in which 16 men with cloacal bladder exstrophy were studied, 14 were transformed into women, two weeks after birth, and two grew up as men. They were between 8 and 21 years old when questioned about their gender identity. In spite of the gender reassignment, only 5 out of 14 had a female identity, 8 adopted a male identity after learning that they had been sexually transfigured. The two persons who did not have sex change remained male and there was no information from one person (BLUMBERG 2009).
The fact that these people, in spite of stunted genitals, feel that they belong to their genetic sex, gives indications that humans are not born gender-neutral.
Intersexuality in the animal kingdom
Incidentally, intersexuality is also documented in the animal kingdom. The geneticist Richard Goldschmidt published in 1931 the work „Die sexuellen Zwischenstufen (The sexual intermediate stages)“ and documented there a number of cases, but mainly in invertebrate animals.
A special form of the intersexual development is the so-called Freemartin-Syndrome and is particularly common in domestic cattle. It is a congenital malformation of females from multiple pregnancy with at least one male fetus. Mammalian embryos are connected to the placenta of the mother’s uterus and thus provide an exchange of substances. If several embryos develop in the womb, the fetal blood vessels of the fetuses can connect to each other, so that it comes to blood exchange. The foreign blood cells are not recognized by the immune system as an attacker, as the fetuses in the womb have a so-called immune tolerance. If we have male and female fetuses in the womb, the sex hormones and the antimatter hormone of the males can also penetrate into the female fetuses. In females, too, an underdevelopment of female sexual organs occurs. Freemartin-Syndrome occurs in 93% of all unequal multiple pregnancies in cattle with males and females in the womb and was described for the first time in 1779 by John Hunter, a Scottish scientist (see BLUMBERG 2009: 223-224, Fig. 2).

Fig. 2: Freemartine cattle in a print from the collection of John Hunter. Source: wikipedia
Freemartine cattle are sterile, but that does not have to be the case with all species. We know from pigs, mice and gerbils, that female fetuses lying between two male fetuses in the uterus also receive high levels of testosterone, which has an impact on their anatomy, physiology and behavior. However, these masculine females are quite fertile if they receive the testosterone levels relatively early in their embryonic development. But not only that: their behavior, including aggression and territorial behavior, can be beneficial for survival and reproduction (BLUMBERG 2009: 230).
A number of intersex individuals have also been observed in brown and black bears. In an adult female brown bear, the clitoris resembled a penis and even had a baculum (penile bone). A female brown bear was found with normally developed fallopian tubes, ovaries, uterus and vagina. At the anterior end of the vagina, however, a penis-like structure with urethra and baculum was found, a vaginal entrance was missing. The penis-like structure corresponded to two-thirds of the usual penis length of male black bears. At the time of her death, the female had two cubs, which means that she had urinated, copulated, and hatched cubs through this penile structure (CATTET 1988).
Heinz-Jürgen Voß’ abolition of the sexes
Already in part three we have critically commented on and refuted some of Heinz-Jürgen Voß‘ theories. Since he is someone with a biological background who, like Butler, wants to „deconstruct“ the sexes, let’s take a closer look at some of his biological „facts“.
Heinz-Jürgen Voß writes about the phenomenon of intersexuality:
By contrast, in the 1920s, Richard Goldschmidt had postulated a „uninterrupted series of sexual intermediates“ and this after the chromosomes X and Y, which were considered important for sex determination, had been found and named several years earlier. Source
It is a pity that Voß does not give us one example of this “ uninterrupted series“ in the article. But sexes cannot be “ uninterrupted „, but are clearly distinguishable as shown. Otherwise, a clear demarcation and distinction as such would not be possible. Besides, what exactly is meant by “ uninterrupted series“? The existence of intersex people proves that there are intermediate stages, but they make up only an extremely small percentage and cannot be defined as clear sex.
Next he writes:
In some mammalian species, the distinction of an X and Y chromosome could not be shown at all. Source
This actually seems to apply to a spiny rat species from Japan. According to a study from the year 2001, representatives of the species Tokudaia osimensis actually do not have sex chromosomes (SOUTO et al., 2001). The Y-associated genes are apparently found in the X-chromosome, both in male and female cell lines. Y-chromosomes are apparently also absent in species of mole voles (genus Ellobius; species Ellobius lutescens and Ellobius tancrei). There is no SRY gene or similar gene either (JUST et al., 2007, MATVEEVSKY et al., 2016, MULUGETA et al., 2016). Their sex determinations (both males and females are XX) are due to differential recombination and X-inactivation during meiosis. However, just because some species of mammals do not have a Y-chromosome, it does not mean that they form different sexes than other mammal species. Both Tokudaia osimensis and Ellobius lutescens and E. tancrei have males and females, nothing in between or a third sex.
In another article, Voß writes:
This often sounds different in popular books and magazines. It often seems that biology knows for sure what „biological sex“ is. But in such articles, biologists, doctors and journalists like to point out that gender is a „sexy topic“ that sells well and protects the research materially and because in the current society only two genders seem dominant. Source
Regardless of the fact that it is certainly a fact that there are research funds for the construction of more than two genders or at least this topic gains attention (after all, it is Voß’ research area on which he lives), biological science has a clear definition of sex. Sexes serve to produce germ cells, of which there are two types, so that two genetic (and epigenetic) systems can unite. It is absolutely outstanding that Voß does not mention this central aspect of sexuality in the biological sense. Much of this article also deals with the critical remark that there is not one sex-determining gene, but that many genes play a role in sex development, and that these genes also have other functions outside of sex determination. This is well-known and has been sufficiently commented on in part three. But that does not change the reality of the sexes, regardless of whether a rat has an X- or Y-chromosome or not! The formation of all other body structures (including metabolism and behavior) are the result of a complex interaction of genes and environment. Our limbs arise from this complex interplay of genes, environment and development (see GILBERT 2006, Chapter 16, pp. 505-528, BLUMBERG 2009, chapters three and four, pp. 105-190). But to deduce that our limbs are social constructs because we divide them into arms and legs is as absurd as to doubt that humans are not tetrapods (possessing four limbs) because there are some who have only one arm!
However, one statement from Voß deserves closer consideration:
„(…) the „male sperm cell“ in the biology and medicine of their time was described as active, flexible and upgraded, whereas the „female egg cell“ was passive, lethargic and in this sense negatively connoted. „Egg“ versus „sperm cell“ represented the place in the society, the woman and man would have to take. The woman is passive, immovable, conservative, not creative – she should therefore not take a central place in politics and science, whereas the man as active, flexible, thinking, changing created just for such positions in society. Thompson countered with such descriptions and made it unmasking that „if one reversed the nature of the genital cells [ovum, sperm cell, note HV], it would be easy for the followers of this doctrine to derive the characteristics of the sex as they did now specify for the opposite case. It would then embody the female cell, smaller and more mobile than the male, the woman with her smaller body size, her excitable nervous system and her inability to attentive attention, while the male cell, large, calm and self-reliant, the size and strength that would represent impartial thinking and the slight concentration of man’s attention.“ (Thompson 1905): Comparative Psychology of the Sexes Experimental Investigations of Normal Mental Abilities in Man and Woman Authorized translation by JE Kötscher, A. Stuber’s Publishing [C. Kabitzsch], Würzburg, p.183)
If you want to say that the androcentric assumptions are a historically distant view, will be disappointed. Rather, considerations that „male gender“ is superior was an important assumption in the development of „modern“ biology and medicine. There are found such descriptions still today. „Female gender“ was described as a basal without any active developmental steps, so it was assumed that „male gender“ would need to have active developmental steps for it to emerge as „superior human form“. When „masculine testicles“ were increasingly recognized as these „masculinizing factors“ in the eighteenth century, established genetics had been searching since the 1940s for a section on chromosomes responsible for the formation of the testicles – the search for the „Testicular determinant factor“. Source
Comment: One can argue without doubt about whether the descriptions of female and male germ cells reflect cultural interpretations. There can really be some truth. But this does not derive from the fact that sex/gender is a “ social construct“. Biology describes real, material, things. There would be no change in the biological significance of the two sexes if the mobile and agile sperm were defined as „female“ and the large, sluggish oocytes as „male“. Such a reversal would not abolish gender discrimination either.
We can only conclude with regard to Voss: he provides no new biological knowledge (at least for biologists, as well as biologically interested), he proves no new sex/genders (even his justified criticism of the cult of genes as the determining factor prove no new sex/genders) and his „queer theory“ on the „deconstruction“ of the sexes/gender is neither scientific (see part one) nor contributes to the end of sex/gender discrimination.
Sex redefined?
In February 2015, a journal article by journalist Claire Ainsworth published a four-page article titled „Sex Redefined“ in the scientific journal Nature, (AINSWORTH 2015).
In this article she essentially mentions factually correct that there are so-called intersex people in addition to male and female individuals. She gives in a table „intermediate forms“ that go from completely male to completely female. In addition to „real“ intersexuals (with chromosome abnormalities), these intermediate stages include those that are genotypically and phenotypically male or female, but are, for example, infertile or have „different genitalia“ (e.g., micropenis). In doing so she does not come to “ uninterrupted series of intermediate forms“, but to 7 intermediate forms + two „correct, fertile“ sexes. These intermediate forms are quite rare. In her section „Beyond the Binary,“ she raises the question of whether have to come out as male or female by law we. Reading the text gives the reader the impression that two-sex is only a variant of a spectrum. On the other hand, even Ainsworth states that 99% of all people are clearly male or female. Ainsworth article evoked critical questions in the research community of why a prestigious journal such as „Nature“ published a gender-critical contribution. KUTSCHERA (2018: pp. 228 – 229) gives a plausible explanation:
The science journal Nature consists of two parts: a front magazine section (according to Kutschera „gossip columns of the natural sciences“) and a scientific section (research papers, peer-reviewed articles, etc.). According to Kutschera, female editors dominate in the „magazine section“, while the „scientific“ section is dominated by men. I would not want to go so far as to separate the two parts by gender (though I do not doubt that men dominate in science). That the magazine section is something like „gossip columns“ may sound pretty rough. However, it seems to be a fact that in addition to scientific publications with hard data, methodology and discussion of results, there is also an area of readers‘ letters, comments and opinions in „Nature“. This also includes Ainsworth article.
About Freya Brown’s „On the Social Construction of Sex“ – Is Gender Science Marxist?
In 2014, „Marxist” author Freya Brown published an article entitled „On the Social Construction of Sex“ (BROWN 2014).
In this article, Brown criticizes the „social construct“ of man and woman.
Brown argues that the medical methods that determine gender (chromosomes, genitals, gonads, hormones, and secondary sexual characteristics) always show some variation, so a division into male and female must be a social construct:
“When we consider all five of these criteria, it becomes clear that a majority of humanity does not fit neatly into the “male” and “female” classifications.”
Brown’s conclusion, however, is not supported by statistical data, she claims something as true, which would have to be proved first. Her point of view is based on their ignorance of how to define gender. Paul COCKSHOTT (2017) sums up Brown’s mistake:
The claim is made that if an infant is classified as female, it must have XX chromosomes, a female sex hormone balance, ovaries, and clearly female genitalia. Expressed in a mathematical formula, this means that Pf represents the probability that an infant is female, Ph indicates the hormones, Pg the genitals, Po the gonads and Pc the sex chromosomes are female. The formula would look like this:
Pf= Ph*Pg*Po*Pc
If we consider that these 4 probability tests do not statistically correlate with each other, i.e. are independent of each other and the probability of born male or female is about 1.1, each P-value can be given as 0.5 (1/2). Entered into the formula this would look like this:
Pf = 0.5 * 0.5 * 0.5 * 0.5 = (0.5)4 = 0.0625 = 1/16
Derived from this formula, 1/16 would be male and 1/16 female, so 1/8 would be assigned to a unique gender. That’s the way it would be, argues BROWN (2014). But there is a significant flaw in Brown’s thinking here: the 5 listed sexual characteristics are not independent! (COCKSHOTT 2017 has omitted the secondary sexual characteristics in his calculation, therefore only 4 are criteria taken, with 5 criteria only 1/32 would be clearly male and female). All 5 features correlate with each other, are dependent on each other and condition each other. For example, the gonads produce sex hormones, which in turn form the sexual characteristics. Important genes are on the chromosomes, though not always on the sex chromosomes. The genes themselves are once again under hormonal control, etc., etc. Consider following example: supposing the infant has two X chromosomes, external female genitalia, produces during puberty more female sexual hormones and secondary sexual characteristics develop, but the ovaries produce none egg cells. The person would be infertile. According to Brown, the person would not be female because one of the sex characteristics is defect. But because all of these features are related, infertile women are just women. No third sex, no social construct, just a woman. The same applies to men and also to the other sexual characteristics! The data of the censuses, the birth certificates as well as the sex determinations by doctors, midwives etc. confirm the correlation of these characteristics. They are not social constructs but describe reality.
The rest of Brown’s article deals with the topic of intersexuality. We have written a lot about this. It would be appropriate to argue that this very small (less than 1%) minority cannot be assigned to a sex/gender. But to generalize from this statistically unrepresentative group contradicts all scientific sampling.
At the end of the article, BROWN (2014) notes that their analysis does not find anything partially Marxist. That’s an understatement to say the least. There is nothing Marxist about BRWON’s analysis. This can be seen only if one looks at the most important works of Marxist theoreticians (e.g. Marx Kapital, Lenin’s analysis of imperialism, etc.) and finds verifiable assertions, statistical material, scientific evidence, footnotes, etc. They use the common scientific methods. These are completely missing in BROWN’s article and they are typical of the social constructivist gender queer left. Unfortunately, this puts “ the Left“ in a bad, dogmatic, ideologically blind, even stupid light. This is shown in KUTSCHERAS (2018) work for example, which sees not only connections between the creationism and the gender ideology, but classifies the latter as „Marxist“, „communist“ etc. This, coupled with a Victorian-era „ideal image of women“, casts Kutschera’s work in a bad light (detailed examples will follow).
Literature
AINSWORTH, B. (2015): Sex redefined. Nature 518, 288-291 https://www.nature.com/polopoly_fs/1.16943!/menu/main/topColumns/topLeftColumn/pdf/518288a.pdf
BLACKLESS, M, CHARUVASTRA, A., DERRYCK, A., FAUSTO-STERLING, A., LAUZANNE, K., LEE, E. (2000): How sexually dimorphic are we? Review and synthesis. Amer. J. Hum. Biol. 12, 151 – 166
BLUMBERG, M. 2009: Freaks of Nature – What Anamolies tell us about Development and Evolution. Oxford University Press
BROWN, F. (2014): On the Social construction of sex https://anti-imperialism.org/2014/02/24/on-the-social-construction-of-sex-part-1/
CATTET, M. (1988): Abnormal sexual differentiation in black bears (Ursus americanus) and brown bears (Ursus arctos). Journal of Mammalogy 69, 849 – 852 https://www.jstor.org/stable/1381646?seq=1#page_scan_tab_contents
COCKSHOTT, P. (2017): Critique of ‘the social construction of sex’ https://paulcockshott.wordpress.com/2017/03/31/critique-of-the-social-construction-of-sex/?fbclid=IwAR2WjMYyMv_ldYP6XXVI8-nvtySk0i2g7-M4BFBV9yjMdxtSoXNdrMec79o
GILBERT, S. (2006): Developmental Biology. 8th Edition. Sinauer Associations
JUST, W et al. (2007): Ellobius lutescens: sex determination and sex chromosome Sex Dev. 2007;1(4):211-21. https://www.ncbi.nlm.nih.gov/pubmed/18391532
KUTSCHERA, U. (2018): Das Gender-Paradoxon – Mann und Frau als evolvierte Menschentypen. LIT-Verlag
MATVEEVSKY et al. (2016): Unique sex chromosome systems in Ellobius: How do male XX chromosomes recombine and undergo pachytene chromatin inactivation? Sci Rep. 2016; 6: 29949. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4947958/
MULUGETA, E. et al. (2016): Genomes of Ellobius species provide insight into the evolutionary dynamics of mammalian sex chromosomes Genome Res. 2016 Sep; 26(9): 1202–1210. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5052041/
SOUTO et al. (2001): Sex determination without the Y chromosome in two Japanese rodents Tokudaia osimensis osimensis and Tokudaia osimensis spp. Mamm Genome. 2001 Jan;12(1):17-21. https://www.ncbi.nlm.nih.gov/pubmed/11178738
WOODHOUSE, C. R. (1998): Sexual function in boys born with exstrophiy, myelomeningocele and micropenis. Urology 52 – 3-11
