
The question of why there are only two sexes (and genders!) has to do with their biological function. This article is about sex, chromosome numbers and germ cell formation.
Sexuality or sexual reproduction is not only exhausted in the multiplication of the number of individuals, but is based on the constant recombination of the parental genetic material. This ensures a certain variability of the organisms. Sex refers to the totality of all phenomena that are put into this service. The biologist August Weismann has already pointed out the origins and significance of sexuality (WEISMANN 1883, 1886). LANE (2009) has dedicated a chapter on the origins and evolution of sexual reproduction; MARGULIS & SAGAN (1986) also wrote a wonderful book (albeit partially elderly). For visually oriented people, I recommend the following video, also the following.
Chromosomes, haploid or diploid?
How is this variability created? Sexuality is the fusion of two cells and their genomes. For this germ cells are produced. It is important to note that the majority of sexually reproducing organisms have a double set of chromosomes. Chromosomes are components of the cell on which the genetic information is stored. However, chromosomes do not only consist of DNA, but also of a number of proteins, mostly so-called histones (Fig. 1). The complex of DNA and proteins is called chromatin. The DNA is thus not free in the cell nucleus, but is in a condensed form. In the case of cell division (mitosis), chromatin, as so-called metaphase chromosomes (metaphase denotes a stage of mitosis), has the densest condensation (compare ALBERTS et al., 2017, CZIHAK 1981, WOLPERT et al., 2007).
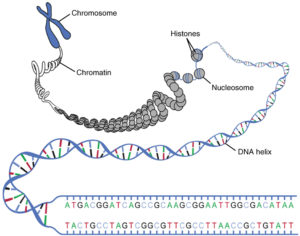
Fig. 1: Condensation of DNA. The DNA is found as a double strand forming a helix. This DNA thread winds around the histones. 4 histones together form a nucleosome. The nucleosomes also wind and form the chromatin framework. The whole is called chromosome. Source: Lecturio
Each species has a certain number of chromosomes. We humans have 46 chromosomes. 44 so-called autosomal chromosomes and 2 sex chromosomes (women: XX, men: XY). In fact, the 46 chromosomes are arranged in 23 pairs. In other words, we have 23 different chromosomes, each of which is duplicated. Double chromosomes are homologous to each other (they are variations of the same chromosome type). A double set of chromosomes is called diploid or 2n. Chromosome sets can be displayed in so-called karyograms (see Fig. 2, see ALBERTS et al., 2017, CZIHAK 1981, WOLPERT et al., 2007).
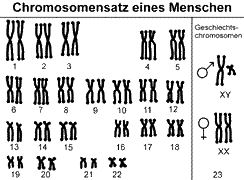
Fig. 2 Karyogram of humans (metaphase chromosomes, in which the genetic material is already duplicated and is distributed in the final phase of mitosis on the two daughter cells). Source
Nerd knowledge: Although our closest relatives, the chimpanzees & bonobos, share 98-99% of our genome, they have 48 instead of 46 chromosomes; so they have one more pair of chromosomes. By the way, for creationists this is the „proof“ that we cannot possibly be related to the monkeys. Obviously, they forget that a research group from the „Chimanzee genome project“ in 2002 proved that our chromosome No. 2 is fused and that in chimpanzees it is split into two chromosomes (2A & 2B). When the ancestors of humans and chimpanzees separated 6 million years ago, our evolutionary line fused the two chromosomes 2A and 2B into one chromosome. So all genes could be preserved. Human chromosome 2 is so similar to chromosome 2A and 2B of the chimpanzee that it had to be a fusion of these. At the site of the fusion there is an about 150,000 base-long part, which is not found in chimpanzees and codes for four genes. Copies of these genes are also found in human chromosome No. 9 (see Fig. 3, DEGROUCHY 1987, FAN et al., 2002).
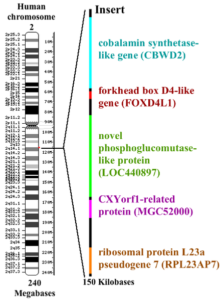
FIG. 3: Fusion of chimpanzee chromosomes 2A & 2B in human chromosome No. 2. The red dot shows the fusion site of both chromosomes to which a 150,000 base pair DNA sequence inserts (shown enlarged to the right). Source: wikipedia
The double set of chromosomes comes about because we have received one half from our father, the other from our mother. Thus, the chromosomes of two different cells have merged to form our double set of chromosomes. Our germ cells, which fuse together, are haploid (1n), so they only have one set of chromosomes. The process in which haploid cells are formed is called meiosis, which is distinguished from actual cell division (mitosis) (see ALBERTS et al., 2017, CZIHAK 1981, WOLPERT et al., 2007).
Mitosis – cell division
Cell division (mitosis, Fig. 4, see ALBERTS et al., 2017, CZIHAK 1981, WOLPERT et al., 2007) takes place in various steps.
Interphase is the process between two cell divisions. In the G1 phase – sometimes called the gap phase – the cell grows and prepares for cell division. There is an increase in protein synthesis. In the S phase, the duplication (replication) of DNA takes place. The G2 phase represents the transition to mitosis; All prerequisites for the core division are now available.
In prophase, the chromosomes contract strongly. They condense and become visible by this process under the light microscope. Each chromosome consists of two identical strands (called chromatids, not to be confused with chromatin!), which largely separate from each other and are held together only by a so-called centromere. At the same time, the formation of the mitosis spindle, a part of the cytoskeleton, which has the task of separating the individual chromatids from each other, begins.
In the metaphase, the chromosomes (consisting of two chromatids = two-chromatide-chromosomes) have shortened even further (strongest condensed form) and arranged in one plane (equatorial plane). The sister chromatids point to the opposite spindle poles. These form fibres (polar microtubules) that attach to the kinetochors of the centromeres.
In the anaphase, the fibres shorten and separate the individual chromatids from each other. In telophase, two diploid cells (2n) are formed, i.e. in humans with 46 chromosomes (= one-chromatid chromosome).
This ensures that the cells receive their respective sets of chromosomes at the time of division.
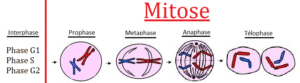
Fig. 4. Mitosis. Source: https://jeretiens.net/difference-entre-mitose-et-meiose/
Meiosis – reduction division
Meiosis, i.e. the formation of haploid (1n) germ cells, proceeds in a similar manner (Fig. 5, compare ALBERTS et al., 2017, CZIHAK 1981, WOLPERT et al., 2007). For a better overview, meiosis is subdivided into different phases, in which, roughly speaking, a first reduction division (meiosis I) and a second equatorial division (meiosis II) are distinguished. Here, too, the DNA is doubled during the interphase. Each chromosome is composed of two sister chromatids containing identical genetic information., which condense in the prophase 1. The chromosomes pair up so that both copies of chromosome 1 are together, both copies of chromosome 2 are together, and so on.
In metaphase 1 of meiosis I, the homologous chromosome pairs now align themselves on both sides of the equatorial plane and one chromosome each (consisting of two chromatids) points to a spindle pole. In mitosis not the homologous chromosome pairs but each individual chromosome arranges on the equatorial plane.
During anaphase 1, the individual chromosomes are pulled to the spindle poles, so during telophase 1, two cells are formed with half the set of chromosomes (1n). However, these are still two-chromatid-chromosomes. In meiosis I the sister chromatids stay together. This is different to what happens in mitosis and meiosis II. Consequently, it comes to the second division (meiosis II), which is similar to mitosis (but in haploid – 1n- cells). In the end, 4 haploid cells (1n) are produced with one-chromatid-chromosomes.
Because the number of chromosomes is reduced in meiosis, it is also called a reduction division.
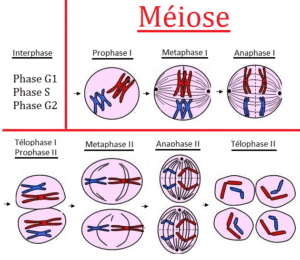
Fig. 5: Meiosis. Source: https://jeretiens.net/difference-entre-mitose-et-meiose/
The meiosis process sounds complicated at first (and there is no need to go more in detail here), but the key aspect we can capture here is the fact that haploid cells are formed during meiosis (1n). These haploid cells are also called germ cells or gametes. A fusion (= fertilization) of two gametes again gives rise to a diploid (2n) cell (the zygote) from which a multicellular organism develops through mitotic division.
Here we already have the first argument, why, biologically speaking, there can be only two sexes: Since we have a double set of chromosomes, we are the product of the fusion of TWO haploid germ cells (gametes).
Biological Life Cycle
Organsimen always have a change between a haploid phase (gametes) and a diploid one. This regular change between a haploid state (haplophase) initiated by meiosis and a diploid state resulting from fertilization (diplophasis) is common to all eukaryotes (cells that have a nucleus, including all animals, plants, fungi, algae and protozoa) that multiply sexually. Depending on which of the two phases predominates, they can be assigned to three different types (Fig. 6, compare ALBERTS et al., 2017, CZIHAK 1981, WOLPERT et al., 2007):
1. Haplontic life cycle
In haplonts, almost the entire development cycle takes place in the haplophase (1n). Only the zygote (= fertilized egg cell) has a diploid chromosome set (2n). This arises from the fusion of two gametes. The resulting zygote then undergoes a reduction division (meiosis). This results in 4 haploid cells again In the whole cycle, zygotes are the only diploid cell; mitosis occurs only in the haploid phase. The haplons include many mushrooms, many algae and some animal protozoa, such as the flagellates.
2. Diplontic life cycle
The diplonts are almost the opposite of the haplonts. With them, the life phase takes place almost exclusively in the diplophase. After fertilization, unlike the haplones, there is no reduction division (meiosis), but the zygote develops into a diploid organism. Some of these cells then form the haploid gametes. In addition to some algae and mushrooms, all multicellular animals (Metazoa) and ciliata (ciliates, unicellular organisms) are among the diplonts.
3. Haplodiplontic life cycle
The haplodiplonts have an intermediate position to the two types mentioned above. They are organisms whose reproduction has alternately a haploid and a diploid generation. This includes most plants. It is characteristic of haplodiplonts that mitoses occur both in the haploid and in the diploid phase. The haploid generation reproduces sexually by forming female and male gametes. It is therefore called gametophyte. The union of two gametes of different sexes and fertilization creates a diploid zygote. The zygote grows to the so-called sporophyte. The sporophytes form haploid spores after a reduction division (meiosis), which reproduce asexually. From these haloid spores, the haploid gametophyte grows and the cycle starts again. In the evolutionary history of plants, the haploid gametophyte is more and more reduced. In mosses the gametophyte is the dominant phase (namely the green moss plant). In ferns, the diploid sporophyte is dominant and in the seed plants, the actual plant is the diploid sporophyte, while the haploid gametophyte is reduced to a few cells (male pollen tube consisting of three cells and female embryo sac in the ovule, consisting of 7-8 cells).
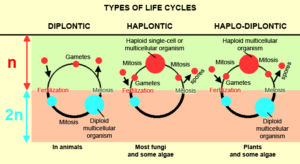
Fig. 6: Biological Life Cycle
Types of germ cells (gametes)
In meiosis, four haploids develop from a diploid cell. But if you look at the gametes, they do not always look the same. The two fertilizing gametes belong to two different types called sexes. Accordingly, there are also two different types of gametes. The smaller or movable gametes are the male gametes, the larger or immobile gametes are called female. The organisms that are able to form the corresponding gametes are classified as male or female. At fertilization, only one gamete of one type can merge with the other. Gametes are therefore the cause of bipolar bipolarity, which results from their functional significance as well as their chemical structure, cell surface, etc. Further morphological differences may be missing in some organisms. Fig. 7 shows the different types of gametes.
If germ cells look morphologically (but not necessarily chemically or functionally!) similar, it is called isogamy. In the case of such isogamy, the two varieties, which are distinguishable only by their functional properties, are arbitrarily designated as + and -, unless a behavioral difference in the initiation of fertilization permits identification of the sex. If the gametes differ in their appearance, this is called anisogamy. If both gametes are flagellated, a distinction is made between male microgametes and female macrogamets. If only one gamete type is flagellated and thus mobile, it is defined as male regardless of its size. If both properties are combined, the small and mobile gametes are the male spermatozoa (called sperm in animals) and the large immobile gametes are the female eggs. This combination is also called Oogamie. It occurs in all animals (including us humans). The reason why oocytes are larger than sperm, is in a special function of meiosis in oocytes. Although they arise from a diploid stem cell, which produces four haploid cells, three of these haploid cells become so-called polar bodies. The division is inequal. From a large cell, only a small cell is separated, which is either dissolved or divides again. This creates a very large egg cell and 3 so-called polar bodies, which are functionless. The production of different gametes naturally goes hand in hand with the formation of different structures of their producers: this applies both to primary sex organs, as well as the secondary ones. In extreme cases, a true sexual dimorphism develops, in which males and females look as different as if they belonged to other species (compare ALBERTS et al 2017, CZIHAK 1981, WOLPERT et al., 2007). Since two different types of gametes per species are always formed in nature, the division into two sexes makes sense here as well. A third gender would only make sense biologically if there are triploid species that emerge from three different germ cells.
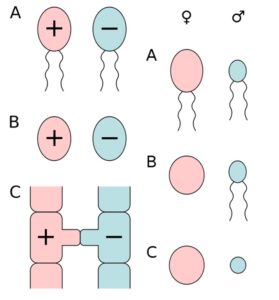
Fig. 7: the different types of gametes. Left column: A) actively moving isogametes, B) stationary isogametes, C) gametangiogamy: fertilization without gametes in the zygomatic fungi. Right column: A) actively moving anisogametes, B) egg cell and sperm (oogamy), C) immovable anisogametes (egg cell and spermatium). Source: wikipedia
Although there are so-called polyploid species, or cells (i.e. their chromosome set is more than double), these are mainly the cause of a faulty meiosis in which the spindle apparatus is not formed or incomplete and so the homologous chromosomes cannot be evenly distributed. So diploid gametes can arise. But here, too, we only have two types of germ cells.
The next parts deal with sex determination, the genetic differences between the sexes, the problem of intersexuality and distress, and the arguments of Fausto-Sterling and Heinz-Jürgen Voß, which are based on more than two genders.
Literature
ALBERTS, B. et. Al (2017): Molekularbiologie der Zelle. Wiley-VCH; Auflage: 6.
CZIHAK, G., LANGER, H., ZIEGLER, H. (1981): Biologie: Ein Lehrbuch (Springer-Lehrbuch). Springer-Verlag, Auflage 2
DEGROUCHY, J. 1987: Chromosome phylogenies of man, great apes, and Old World monkeys. Genetica. 1987 Aug 31;73(1-2):37-52. https://www.ncbi.nlm.nih.gov/pubmed/3333352
FAN, Y., NEWMAN, T., LINDARDOPOULOU, E. & TRASK, B. J. (2002): Gene Content and Function of the Ancestral Chromosome Fusion Site in Human Chromosome 2q13–2q14.1 and Paralogous Regions. Genome Res. 2002 Nov; 12(11): 1663–1672. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC187549/
LANE, N. 2009: Life Ascending: The Ten Great Inventions of Evolution. Profile Books
MARGULIS, L. & SAGAN, D. 1986: Origins of Sex: Three Billion Years of Genetic Recombination. Yale University Press
WEISMANN, A. 1883: Die Entstehung der Sexualzellen bei Hydromedusen. Zugleich ein Beitrag zur Kenntnis des Baues und der Lebenserscheinungen dieser Gruppe. Verlag Gustav Fischer
WEISMANN, A. 1886: Die Bedeutung der sexuellen Fortpflanzung für die Selektionstheorie. Verlag Gustav Fischer
WOLPERT, L et al. (2007): Principles of Development: Das Original mit Übersetzungshilfen: Das Original Mit Übersetzungshilfen
Video-Beiträge:
The Origin of Sexual Reproduction
