
Hallo zusammen, vielen Dank für die Kommentare und Themenvorschläge in meinem letzten Dinosaurier-Video zu welchen Merkmalen der Evolution der Dinosaurier zu den modernen Vögeln ich Videos machen soll. Es wurden viele tolle Vorschläge gemacht und zu den meisten wird sicherlich etwas kommen. Doch in diesem Video bleibe ich tatsächlich noch im Hüftbereich und wir schauen uns das Becken der Dinosaurier und Vögel etwas genauer an.
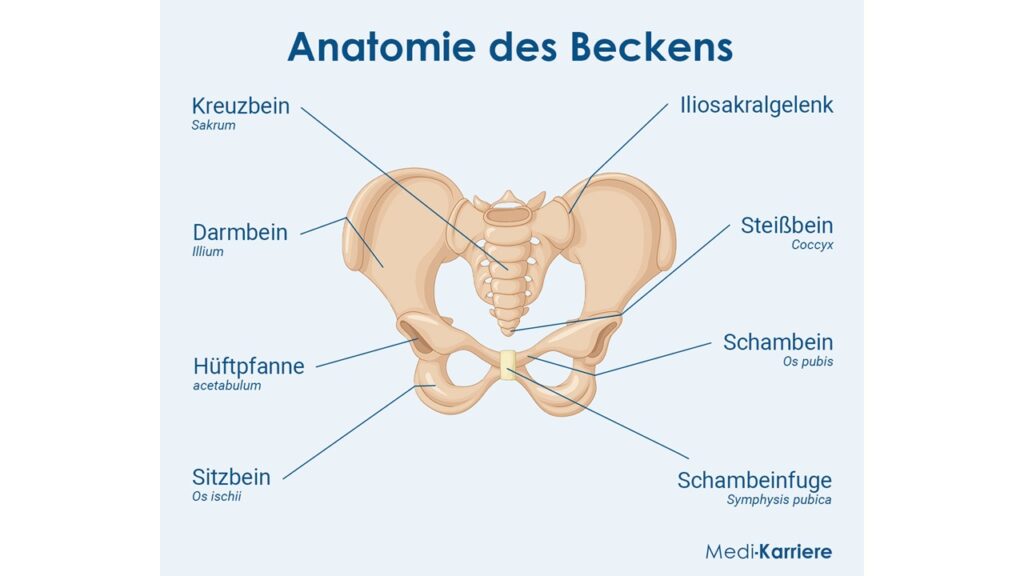
Aus was besteht ein Becken?
Ein jedes Wirbeltierbecken besteht aus drei verschiedenen Knochen pro Körperseite, die mit dem Kreuzbein verbunden sind. Betrachten wir dafür das Becken eines Menschen. Zum einen haben wir das Darmbein oder Ilium. Beim Menschen fallen vor allem die großen Darmbeinschaufeln auf. Sie dienen primär als Ansatzstellen für viele Muskeln. Während das Darmbein die vordere Stabilitätssäule des Beckens bildet, wird die hintere durch das Sitzbein, auch als Ischium bezeichnet, gebildet. Wie der Name verkündet, ist dies der Knochen, auf dem wir sitzen. Der dritte Teil des Hüftgelenks ist das Schambein, auch Pubis genannt, welches sich vorne unten befindet. Beide Schambeinknochen werden im vorderen Bereich durch die Schambeinfuge verbunden, welche so den Beckenring verschmelzen. Das größte Gelenk im Bereich des Beckens ist das Hüftgelenk, dessen Gelenkpfanne, das Acetabulum, von allen drei Hüftknochen gebildet wird. In ihr inseriert der Oberschenkelkopf, also der oberste Teil des Oberschenkelknochens.
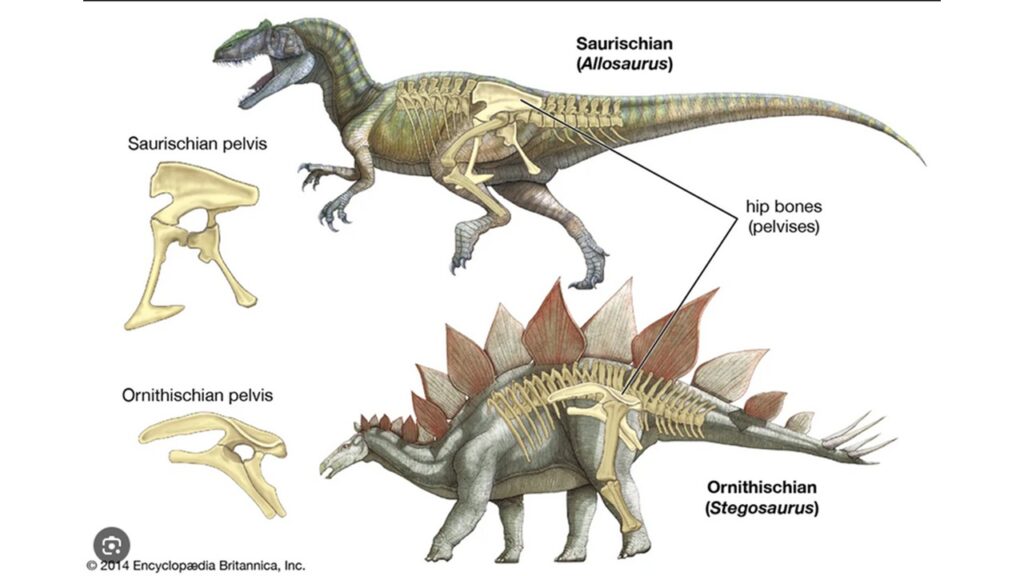
Beckenknochen der Dinosaurier
Diese grundlegende Muster aus Darmbein, Sitzbein und Schambein findet sich auch bei den Dinosauriern, doch ihre Form unterscheidet sich von jenen der Menschen. Ein charakteristisches Merkmal der Dinosaurierbecken bildet die Hüftgelenkpfanne, das Acetabulum. Während das Acetabulum des Menschen und vieler Landwirbeltiere knöchern ist, findet sich im Dinosaurierbecken eine Öffnung, die direkt durch die Hüftknochen geht. Das heißt Dinosaurier hatten ein offenes Acetabulum, war also nicht knöchern, sondern höchstwahrscheinlich knorpelig. Ein offenes Acetabulum ist ein diagnostisches Merkmal der Dinosaurier und findet sich auch bei den modernen Vögeln. Nur bei einigen später entwickelten Dinosauriergruppen wie den Ankylosauriern wurde das Acetabulum wieder knöchern.
Die verschiedenen Dinosauriergruppen werden dann anhand der Form der Beckenknochen in zwei Hauptgruppen unterschieden. Bei den Echsenbecken-Dinosauriern oder Saurischia ist das Schambein nach vorne gerichtet. Dieses Merkmal findet sich bei den langhalsigen Sauropoden und den fleischfressenden Theropoden. Besonders beim Tyrannosaurus ist das nach vorne gerichtete Schambein gut zu erkennen. Die restlichen Dinosaurier werden, dann als Vogelbecken-Dinosaurier oder Ornithischia bezeichnet, bei denen ein Teil des Schambeins nach hinten ragt und parallel zum Sitzbein verläuft. Zu den Vogelbecken-Dinosauriern gehören die Ankylosaurier, Horndinosaurier, die Entenschnabeldinosaurier und alle weiteren Pflanzenfressenden Dinosaurier außerhalb der Sauropoden. Und hier stehen wir vor einer interessanten Ironie: Vögel haben ein nach hinten gerichtetes Schambein, welches sich aber in den Details von jenen der Vogelbeckendinosaurier unterscheidet. Aufgrund dieser Ähnlichkeit mit dem Becken der Vögel wurden diese Dinosaurier als Vogelbecken-Dinosaurier bezeichnet. Aber: Vögel sind eine Untergruppe der Theropoden, gehören also zu den Echsenbecken-Dinosauriern. Das heißt das Vogel-Becken entstand aus einem Echsenbecken und nicht aus dem Becken der Vogelbecken-Dinosaurier. Wenn das zutrifft, dann müssen wir im Fossilbericht sehen können, wie einzelne Theropoden-Dinosaurier mit einem Echsenbecken mit zunehmender Verwandtschaft ein immer Vogel-ähnlicheres Becken haben müssen, bei dem das Schambein nach hinten gerichtet ist. Genau das liegt vor.
Bei den Maniraptoren, die eng mit den Vögeln verwandt sind, reicht das Schambein weiter nach hinten. Es zeigt sich also eine zunehmende Verlagerung des Schambeins nach Hinten in die „vogelartige“ Position. Diese schrittweise Umwandlung ist durch einen hervorragenden Fossilnachweis gut dokumentiert.
Embryonalentwicklung der Beckenknochen
Diese Umwandlung des Beckens ist aber nicht nur durch den Fossilbericht dokumentiert, sondern auch in der Embryonalentwicklung.
In neuen Forschungsarbeiten haben Wissenschaftler der Universität Yale embryologische Bildgebungsverfahren eingesetzt, um das Beckengewebe von Vögeln in 3D zu untersuchen, was einen direkten Vergleich mit dem Fossilnachweis ermöglicht. In ihrer Studie untersuchten sie die Entwicklung des Beckens bei Alligatoren, Haushühnern, japanischen Wachteln, chilenischen Steißhühnern und Sittichen und verglichen ihre Entwicklungsstadien mit denen von Dinosauriern, einschließlich des Archaeopteryx. Sie markierten embryonale Becken mit Antikörpern, um nach Proteinen zu suchen, die in sich entwickelnden Knorpeln, Bindegewebe, Skelettmuskeln und Nerven exprimiert werden. Anschließend erstellten die Forscher mit Mikroskopen und CT-Scans 3D-Bilder des Beckens, der Muskeln und der Nerven. Dabei zeigte sich etwas Bemerkenswertes: Viele Merkmale der Dinosaurier-Vorfahren der Vögel treten vorübergehend in der frühen Embryonalentwicklung auf. Dazu gehört zum Beispiel ein nach vorne gerichtetes Schambein – ein typisches Merkmal vieler Dinosaurier. Erst im Verlauf der Entwicklung nimmt das Becken seine typische vogelartige Form an. Die Embryonalentwicklung durchläuft dabei eine Abfolge von Veränderungen, die stark an die evolutionäre Abfolge dieser Merkmale erinnert. Vereinfacht gesagt: Jeder Vogel beginnt seine Entwicklung mit einem „dinosaurierähnlichen“ Becken.
Warum haben Vögel ein offenes Becken – und Dinosaurier nicht?
Schauen wir uns ein weiteres Merkmal des Beckens genauer an: die Schambeinfuge, fachlich Symphysis pubica. Das ist eine knorpelige, kaum bewegliche Verbindung, die auf der Vorderseite des Beckens das rechte und linke Schambein miteinander verbindet.
Die meisten Wirbeltiere besitzen eine solche Schambeinfuge. Vögel bilden hier jedoch eine bemerkenswerte Ausnahme. Bei fast allen Vogelarten ist das Becken „offen“ – die beiden Schambeine treffen sich also nicht. Unter den heute lebenden Vögeln ist der Strauß meines Wissens die einzige Art, bei der noch eine Schambeinfuge vorhanden ist.
Bei Dinosauriern hingegen war diese Verbindung üblich, auch bei den gefiederten Nicht-Vogel-Maniraptoren. Ebenso besitzen alle mesozoischen Avialae – darunter Archaeopteryx, Confuciusornis, Hesperornis sowie verschiedene Enantiornithes – noch eine Schambeinfuge. Auch Gansus, ein Vogel, der den modernen Vögeln bereits sehr nahe steht, zeigt dieses Merkmal.
Das deutet darauf hin, dass die Schambeinfuge erst innerhalb der Linie der modernen Vögel verloren ging – wahrscheinlich irgendwo innerhalb der Ornithurae und schließlich bei den heutigen Vögeln, den Aves.
Der Strauß steht an der ursprünglichsten Abzweigung innerhalb der modernen Vögel. Daher gibt es zwei mögliche Erklärungen: Entweder ging die Schambeinfuge erst innerhalb der Aves verloren – oder sie verschwand bereits früher und wurde beim Strauß sekundär wieder geschlossen. Aber warum besitzen moderne Vögel überhaupt ein offenes Becken?
Das Fehlen einer Schambeinfuge ermöglicht es Vögeln, verhältnismäßig größere Eier zu legen.
Bei mesozoischen Vögeln mit Schambeinfuge war der Raum begrenzt, durch den ein Ei den Körper verlassen konnte. Studien zeigen daher, dass diese ausgestorbenen Vögel – im Verhältnis zu ihrer Körpermasse und zur Größe der Schambeinfuge – kleinere Eier legten als moderne Vögel. Wichtig ist dabei: Es geht nicht um die absolute Größe eines Eis, sondern um seine Größe im Verhältnis zum erwachsenen Tier.
Relativ größere Eier bieten mehrere Vorteile. Sie sind stabiler und besser geeignet für das sogenannte Kontaktbrüten, also wenn sich das Elterntier direkt auf die Eier setzt, um sie zu wärmen.
In Zeiten von Umweltkrisen oder Massenaussterben kann das ein entscheidender Vorteil sein: Die Eier sind besser geschützt und werden effizienter bebrütet. Dadurch steigen die Überlebenschancen der Küken – und damit auch mehr Nachkommen und größere evolutionäre Fitness.
Diese Erkenntnis ist insofern interessant, als dass für Kreationisten Archaeopteryx, Confuciusornis und andere mesozoische Vögel, nur Vögel und keine Dinosaurier sind. Aber diese mesozischen Vögel teilen wichtige Merkmale mit den Dinosauriern – so die Schambeinfuge. Kreationisten beginnen mit ihrer Kindergarten-Taxonomie so den typischen Kardinalsfehler.
Microraptor: Dinosaurier oder Vogel?
Und das bringt mich nun zum letzten heutigen Punkt: Microraptor. Im Stammbaum der Dinosaurier gehört Microraptor zu den Dromeosauridae, ist also eng mit Velociraptor und Deinonychus verwandt. Damit ist er zwar mit den Vögeln eng verwandt, steht aber außerhalb der Avialae, zu der Archaeopteryx, Jeholornis, Sapeornis, Confuciusornis, die Enantiornithes, Ganus, Hesperornis, Ichthyornis und die modernen Vögel gehören. Er ist also, salopp formuliert, mehr Nicht-Vogel-Dinosaurier als Vogel, weist aber eine Reihe charakteristischer Merkmale auf, die er (und andere Raubsaurier) mit Vögeln teilen, z. b. das Vorhandensein von Federn. Interessanterweise ist für Kreationisten, wie den Irren von Answers in Genesis, Microraptor genau deswegen ein Vogel: er hat Federn und alles was Federn hat, ist für sie ein Vogel und kein Dinosaurier. Dabei werden alle anderen morphologischen Nuancen bewusst ignoriert. In einem Artikel versucht Answers in Genesis zu beweisen, dass Microraptor ein Vogel und kein Dinosaurier sei. Sie diskutieren dabei viele Merkmale und zitieren aus dem Zusammenhang gerissene Studien, längst widerlegte Einzelmeinungen und ignorieren sämtliche Gegenteilige Befunde. Konzentrieren wir uns hierbei auf ein kleines Merkmal: den supracetabularen Kamm. Kompliziertes Wort? Dann kurz erklärt: das Acetabulum ist, wie zuvor erwähnt, die Hüftgelenkpfanne, die den Oberschenkelkopf aufnimmt und so das Hüftgelenk bildet. Ein supraacetabulärer Kamm ist eine knöcherne Leiste am Darmbein oberhalb der Hüftgelenkpfanne, der das Becken stützt und so eine aufrechtere Körperhaltung ermöglicht. Dieser Kamm fehlt bei modernen Vögeln, ist aber bei Nicht-Vogel-Dinosauriern vorhanden. Answers in Genesis behauptet nun, Microraptor hätte diesen Kamm nicht und sei somit ein Vogel und kein Dinosaurier. Sie ignorieren jedoch die Tatsache, dass eine spätere und detailliertere Analyse von Microraptor gezeigt hat, dass solch ein Kamm bei Microraptor vorhanden ist. Der Grund, warum er bei der Erstbeschreibung als fehlend angegeben wurde, war, dass das Becken zerquetscht und so deformiert war. Darüber hinaus war Microraptor eng mit dem kürzlich beschriebenen gefiederten Dinosaurier Hesperonychus verwandt, dessen Becken unzerquetscht erhalten geblieben ist. Bei diesem Dinosaurier ist ein supraacetebulärer Kamm vorhanden. Aber unabhängig davon, verstrickt sich Answers in Genesis in weitere Widersprüche. Denn bei Archaeopteryx, Anchiornis, Aurornis, Xiaotingia, Velociraptor usw., die von Answers in Genesis alle als Vögel und nicht als Dinosaurier bezeichnet werden ist ein markanter supraacetabulärer Kamm dokumentiert. Wie immer widerlegen Kreationisten sich selbst. Nach ihrer eigenen Aussage ist dieses Merkmal ein diagnostisches Merkmal von Dinosauriern und finde sich nicht bei Vögeln. Doch genau diesen Kamm finden wir bei ursprünglichen Vögeln, die auch von Kreationisten als Vogel anerkannt werden. Man macht sich die Welt wie sie einem gefällt. Es ist fast so, als wären die von uns als Vögel bezeichneten Tiere in Wirklichkeit nur kleine gefiederte Dinosaurier. Und Federn sind doch das, was einen Vogel so vogelartig macht. Und genau das Merkmal schauen wir uns im nächsten Video an. Bliebt also dran für noch spannenderen Dino-Content.
Literatur
Baron, MG, Williams, ME (2018). A re-evaluation of the enigmatic dinosauriform Caseosaurus crosbyensis from the Late Triassic of Texas, USA and its implications for early dinosaur evolution. Acta Palaeontologica Polonica 63 (1): 129–145. https://doi.org/10.4202/app.00372.2017
Bindellini, G et al. (2025). Velociraptor : the state of the art 100 years after the discovery of this iconic dinosaur. Italian Journal of Geosciences 144: 460-486 https://doi.org/10.3301/IJG.2025.22
Brougham, J, Brusatte, SL (2010). Distorted Microraptor specimen is not ideal for understanding the origin of avian flight, Proc. Natl. Acad. Sci. U.S.A. 107 (40) E155-E155, https://doi.org/10.1073/pnas.1004977107.
Charles Deeming D, Mayr, G (2018). Pelvis morphology suggests that early Mesozoic birds were too heavy to contact incubate their eggs. J. Evol. Biol., 31: 701-709. https://doi.org/10.1111/jeb.13256
Griffin, CT et al. (2022). The developing bird pelvis passes through ancestral dinosaurian conditions. Nature 608, 346–352. https://doi.org/10.1038/s41586-022-04982-w
Longrich, NR, Currie, PJ (2009). A microraptorine (Dinosauria–Dromaeosauridae) from the Late Cretaceous of North America, Proc. Natl. Acad. Sci. U.S.A. 106 (13) 5002-5007, https://doi.org/10.1073/pnas.0811664106.
Rhodes MM, Henderson DM, Currie PJ. (2021). Maniraptoran pelvic musculature highlights evolutionary patterns in theropod locomotion on the line to birds. PeerJ 9:e10855 https://doi.org/10.7717/peerj.10855
Xu, X et al. (2011). An Archaeopteryx-like theropod from China and the origin of Avialae. Nature 475, 465–470. https://doi.org/10.1038/nature10288
