

Betritt man ein Naturkundemuseum fallen sie einem sofort ins Auge: Fossilien. Am bekanntesten und beliebtesten sind natürlich die Dinosaurier, Eiszeittiere wie Säbelzahnkatzen und Mammuts und natürlich unsere eigenen Vorfahren. Doch was sind Fossilien überhaupt? Wie entstehen sie und was kann man aus ihnen herauslesen? Diese Fragen wollen wir hier beantworten.
Was ist ein Fossil?
(Text als Pdf)
Grundsätzlich: Fossilien sind Überreste ausgestorbener Lebewesen, die vor mindestens 10.000 Jahren gestorben sind.
Dass ein Lebewesen zum Fossil wird, ist ein sehr unwahrscheinliches Ereignis. Die meisten Lebewesen werden vorher gefressen oder verwesen, bevor sie eine Möglichkeit haben zu fossilisieren. Dass Fossilien entstehen können, bedarf ganz besonderer Bedingungen. Dennoch konnten Wissenschaftler bis 1999 etwa 250.000 fossile Tier- und Pflanzenarten wissenschaftlich beschreiben (Prothero 1999).
Fossilien findet man auch nicht in allen Gesteinsschichten, sondern nur in Sedimentgesteinen. Geologen unterscheiden drei Typen des Gesteins: magmatische Gesteine, Sedimentgesteine und metamorphe Gesteine Abb. 1). Sedimentgesteine im geowissenschaftlichen Sinn sind verschiedene mineralische (anorganische) und/oder organische Lockermaterialien, die – nach einem kürzeren oder längeren Transport durch Schwerkraft oder ein strömendes Medium – auf dem trockenen Land oder am Grund eines Gewässers abgelagert werden.
Magmatische Gesteine sind Steine, die durch vulkanische Einwirkung entstanden sind und metamorphe Gesteine, sind welche, die sich tief in der Erdkruste durch die Metamorphose anderer Gesteinsarten gebildet haben.
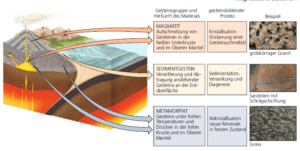
Abb. 1: Gesteinsarten
Tiere, die normalerweise in Sedimenten leben, werden eher im Sediment begraben, bevor sie zerstört werden, und Arten, die auf der Oberfläche des Sediments (d. h. auf dem Meeresboden) leben, versteinern eher als Arten, die im freien Wasser schwimmen. An Land lebende Arten werden am seltensten zum Fossil. Das mag einen verwunderlich klingen, denn die für den Nichtfachmann bekanntesten Fossilien sind interessanterweise die Landtiere, wie Dinosaurier, Eizeittiere oder unsere Vorfahren. Die häufigsten Fossilien findet man bei Korallen, Schwämmen, Trilobiten, Krebstieren, Ammoniten und den Foraminiferen. Je weiter eine Art von Sedimenten lebt, desto geringer ist die Wahrscheinlichkeit, dass sie versteinert.
Wie entsteht ein Fossil?
Als allererstes muss ein Lebewesen sterben. Seine sterblichen Überreste müssen aber an einem Ort zu liegen kommen, wo sie nicht vollständig von Aasfressern oder durch Verwitterung zersetzt werden. Das kann zum Beispiel begraben unter Sand, Schlamm oder Lehm der Fall sein. Ein guter Ort dafür ist zum Beispiel ein Flussbett oder der Meeresgrund (Abb. 2).

Abb. 2: Entstehung eines Fossils
Die Einbettung in Sedimente verhindert, dass Luft zum Körper gelangt, und verlangsamt so die Verwesung. Im Laufe der Zeit wird der tote Körper immer tiefer begraben, da sich neue Sedimentschichten auf ihn legen. Durch den so entstehenden Druck wird Schlamm und Sand langsam zu Stein zusammengepresst. Der steigende Druck wirkt sich auch auf die eingegrabenen Reste der Lebewesen aus. Diese werden dadurch zusammengedrückt und alles Wasser aus ihnen herausgepresst. Hier kann es passieren, dass Teile des Körpers auch zerstört oder verformt werden. Wenn sich die Sedimente verdichten, werden sie allmählich zu Sedimentgestein. Anschließend können sie durch tektonische Bewegungen der Erdplatten nach oben, unten oder um den Globus bewegt und wieder freigelegt werden.
Im Laufe der geologischen Zeit werden die ursprünglichen harten Teile eines Organismus im Sedimentgestein liegend umgewandelt. Mineralien aus dem umgebenden Gestein imprägnieren langsam die Knochen oder die Schale des Fossils und verändern seine chemische Zusammensetzung. Man spricht von Umkristallisation.
Die weichen Teile eines Körpers – Muskeln, Fett, Organe etc. – überstehen die Umwandlung zum Fossil nur in äußerst seltenen Fällen. Es gibt aber einige Fossillagerstätten, die besonders gut erhaltene und vollständige Fossilien beinhalten: inklusive Köperumriss, Mageninhalt und Weichgewebe. Die Funde der Grube Messel sind hierfür ein gutes Beispiel.
Als Fossilien gelten übrigens nicht nur die Überreste des Lebewesens selbst, sondern auch die Spuren, die ein Fossil hinterlassen kann: sogenannte Spurenfossilien. Am bekanntesten sind natürlich die Fußspuren, die ein Lebewesen hinterlassen kann. Erwähnenswert sind hier z. B. die Dinosaurierfußspuren, die man in Münchehagen, Niedersachen, gefunden hat: über 250 Abdrücke von großen langhalsigen Sauropoden. Berühmt sind auch die Fußspuren von drei aufrecht gehenden Frühmenschen in Laetoli, Tansania (Abb. 3 und 4).

Abb. 3: Dinosaurierfußspuren in Münchehagen

Abb. 4: Hominiden Fußspuren von Laetoli
Der Erhalt von Laufspuren setzt voraus, dass das sie tragende Sediment und das sich auflagernde Sediment aus verschiedenen Substraten bestehen, da sonst die beiden Schichten untrennbar miteinander verschmelzen und zu einer strukturlosen Schicht werden. Häufig sind feuchte Sandufer mit Spuren von Tieren, die zur Tränke kamen. Der Sand wurde kurz darauf langsam von Wasser überspült, wobei sich Flusssediment absetzte.
Die Auswertung von Laufspuren ist sehr effektiv, da man aufgrund moderner Erkenntnisse der Bewegungsphysiologie, aber auch der Sportwissenschaft sehr genau weiß, wie welche Formen von Abdrücken unter welchen Umständen zustande kommen. So sind Rückschlüsse möglich auf das Gewicht des Tiers, die Laufgeschwindigkeit, das Lebensalter, den Beckenbau bis hin zu etwaigen Verletzungen.
Weitere Spurenfossilien sind Grabspuren, Kriechspuren, Fressspuren oder Kotspuren. Versteinerter Kot wird als Koprolith bezeichnet (Abb. 5). Spurenfossilien werden auch Ichnofossilien genannt.

Abb. 5: Koprolith eines pflanzenfressenden Dinosauriers
Ein weiterer Fossilisationsprozess ist die Inkohlung (Abb. 6). Hierbei findet unter Luftabschluss eine Umwandlung des organischen Materials statt, bei der vorwiegend die Elemente Sauerstoff, Wasserstoff und Stickstoff entfernt werden, wodurch sich der Kohlenstoff relativ anreichert, bis fast nur noch Kohlenstoff übrigbleibt. Dabei können mit zunehmender Inkohlung Braunkohle oder Steinkohle entstehen. Der Prozess kommt bei Pflanzenmaterial vor. Das meiste heute geförderte Erdöl ist aus abgestorbenen Meereskleinstlebewesen entstanden, wobei Algen den mit Abstand größten Anteil an Biomasse gestellt haben. Fossile Energieträger haben daher ihren Namen: wir verbrennen wortwörtlich Fossilien!
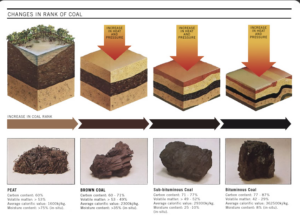
Abb. 6: Inkohlung
Eine weitere Form der Fossilisation wurde durch den Film Jurassic Park berühmt: Bernstein (Abb. 7). Bernstein ist fossiler Baumharz. Einschlüsse wie z.B. Insekten, Spinnen, Blätter oder sogar Kriechtiere sind hier möglich. Besonders aufsehenerregend war der 99 Mio. Jahre alte Fund eines Kükens aus der Vogelgruppe der Enantiornithes in Bernstein! Dinosaurier klonen wird man aber mittels in Bernstein eingeschlossener Blutsauger nicht können. Die meisten Bernsteine sind erdgeschichtlich jünger und die Wahrscheinlichkeit, dass eine Mücke kurz vor ihrem Einschluss in Baumharz eines jedermann Lieblings-Dino gestochen hat und das Blut nicht verdaut wurde, ist sehr unwahrscheinlich.

Abb. 7: Bernsteinfossil
Als letzte Fossilisationsart sei der Steinkern erwähnt. Steinkerne entstehen aus wirbellosen Meerestieren mit Außenskelett oder ähnlichen, gehäuseartigen Hartteilen, wie wir sie z. B. von Muscheln kennen. Diese Hartteile bestehen in der Regel aus Kalziumkarbonat und haben ein deutlich höheres Erhaltungspotenzial als der Weichkörper im Inneren des Gehäuses. Wenn dieser nach der Einbettung vollständig zersetzt ist, bleibt ein hohles Gehäuse im Sediment zurück. Dieses kann nachfolgend mit Sediment verfüllt werden. Wird nach der Verfüllung das eigentliche Gehäuse infolge zerstört oder aufgelöst, bildet dessen nunmehr zu Gestein verfestigte Sedimentfüllung das Gehäuse nach wie vor als Ausguss ab.
Fossilienpräparation
Das Freilegen von Fossilien ist eine mühevolle Arbeit – und der Fund ist erst der Anfang. Meistens ragt nur ein winziges Stück eines versteinerten Knochens aus einem Steinbrocken heraus. Was sich genau darin verbirgt, zeigt sich erst, wenn das umgebende Fremdgestein entfernt ist. Paläontologen nutzen die verschiedensten Werkzeuge um die Fossilien freizulegen – auch abhängig vom Sedimentgestein. Neben Hammer und Meißel, sind vor allem Präparationswerkzeuge wichtig, die man vom Zahnarzt kennt. Damit kann man z. B. Sandstein vorsichtig vom Fossil entfernen.
Wichtig ist auch ein Druckluft-Stichel, ähnlich wie er auch beim Zahnarzt zum Einsatz kommt. Durch die schnellen Stöße eines kleinen Meißels löst sich das Gestein am Rand des Fossils und splittert ab. Eine mühsame Arbeit: Es kann ein ganzes Jahr vergehen, bis ein Präparator ein kompliziertes Stück, wie zum Beispiel einen Schädelknochen, vollständig freigelegt hat. Viele Museen kommen mit dem Präparieren daher kaum noch nach. In vielen Museumskellern lagern noch Tonnen an Gesteinsmaterial, was noch Arbeit für viele Jahre, wenn nicht sogar Jahrzehnte bedeutet.
Andere Fossilien erfordern besondere Methoden. So z. B. die Fossilien der Grube Messel bei Darmstadt. Die Ölschiefer des ehemaligen Sees bergen einen solchen Reichtum an einzigartigen Funden, dass dieses Naturdenkmal mittlerweile zum Weltnaturerbe erklärt wurde. Berühmt ist die Grube Messel vor allem für seine Urpferdchen. Aufgrund der Vergänglichkeit des Ölschiefers (Ölschiefer lässt sich nicht konservieren und zerfällt beim Austrocknen), werden alle Messelfossilien mit der sog. Kunststofftransfermethode präpariert. Dabei wird zuerst eine Seite des Fossils freigelegt und anschließend eine Schicht aus Kunststoff aufgetragen. Nach dem Aushärten wird das Fossil von der anderen Seite freigelegt und der Ölschiefer vollständig entfernt. Durch diese Methode, die von allen wissenschaftlichen Instituten bei den Fossilien aus der Grube Messel angewandt wird, werden die wertvollen Stücke dauerhaft konserviert.

Abb. 8: Präparationswerkzeug
Fossilien studieren
Fossilien sind Zeugnisse des vergangenen Lebens auf unserer Erde. Dieses vergangene Leben zu studieren ist die Aufgabe der Paläontologie. Sie hat als Ziel die Fossilien als ehemals lebendige Wesen zu verstehen, die atmeten, sich ernährten, sich fortpflanzten und starben, in einer längst vergangenen, aber realen Zeit, die wir weder direkt sehen, fühlen oder riechen können. Hieraus ergibt sich natürlich ein Problem, vor welchem z. B. auch Archäologen stehen: Woher wissen wir, was in der Vergangenheit wirklich geschehen ist, ohne daraus eine ausgedachte Geschichte zu machen?
Alles, was wir über die Biologie ausgestorbener Organismen vermuten, sollte im Hinblick auf das, was wir über die Biologie heute lebender Organismen wissen, Sinn machen, es sei denn, es gibt sehr gut fundierte Indizien, die gegenteiliges schlussfolgern lassen. Diese Regel gilt für die gesamte Biologie, von der Zellbiochemie bis hin zu Genetik, Physiologie, Ökologie, Verhalten und Evolution. Da Fossilien in Gesteinen gefunden werden, haben wir Zugang zu Umweltinformationen über den Lebensraum des ausgestorbenen Organismus: Ein Gestein könnte beispielsweise eindeutige Hinweise darauf aufweisen, dass es unter Wüstenbedingungen oder an einem Flachwasserriff abgelagert wurde. Fossilien sind also keine isolierten Objekte, sondern Teile eines größeren Puzzles. Zum Beispiel ist es schwierig, die Biologie der ersten Landtiere zu interpretieren, wenn wir nicht die Indizien berücksichtigen, die in den devonischen Sedimenten erhalten sind, in denen sie aufbewahrt werden und auf die damaligen Umweltbedingungen Rückschlüsse liefern.
Man sollte in der Lage sein, drei Ebenen der paläontologischen Interpretation zu erkennen. Dies wollen wir anhand einer ausgestorbenen Gruppe versuchen verständlich zu machen: der Ichthyosaurier.
Erstens gibt es unvermeidliche Schlussfolgerungen, für die es keine möglichen Alternativen gibt. Es besteht zum Beispiel kein Zweifel daran, dass Ichthyosaurier schwimmende Meeresreptilien waren.
Die zweite Ebene ist die der wahrscheinlichen Interpretation. Hier kann es mehrere Alternativen geben, aber eine große Anzahl der Indizien stützt eine führende Interpretation. Zum Beispiel gibt es gute Belege dafür, dass Ichthyosaurier lebende Junge zur Welt brachten, anstatt Eier zu legen. Fast alle Paläontologen halten dies für die beste Hypothese und wären überrascht, wenn gegenteilige Belege auftauchen würden.
Die dritte Ebene ist die der Spekulationen. Diese können schlüssig sein und stimmen, aber es gibt nicht viele wirkliche Belege für die eine oder andere Spekulation. Paläontologen dürfen Spekulationen als vorläufige Ideen akzeptieren, mit denen sie sorgfältig arbeiten und testen können, aber sie sollten nicht überrascht oder verärgert sein, wenn sich diese als falsch erweisen, wenn sie durch weitere Belege widerlegt werden. Es erscheint zum Beispiel vernünftig, dass Ichthyosaurier gleichwarme Tiere waren, aber es ist eine spekulative Idee, weil sie schwer zu testen ist. Wenn neue Belege zeigen, dass die Idee unwahrscheinlich ist, wäre man persönlich vielleicht enttäuscht, aber wissenschaftlich nicht beunruhigt.
Für Studenten der Paläontologie ist es aber wichtig nicht in die Apologetik zu verfallen und zu sagen: „Nun das geschah vor Millionen von Jahren, also können wir nur spekulieren!“ Das stimmt so nicht. Wir können nämlich die Informationen aus verschiedenen unabhängigen Quellen bekommen und unsere Hypothesen mit anderen, unabhängigen Datensätzen vergleichen. Wir haben z. B. im Beitrag über das Alter der Erde festgestellt, dass das Erdalter mit verschiedenen Methoden getestet werden kann, von der radiometrischen Datierung, über der Auszählung von Tagesringen pro Jahr bei fossilen Korallen und Muscheln, der Ermittlung der Geschwindigkeit der Kontinentaldrift, der Auswertung des kosmischen Mikrowellenhintergrundes usw. Nicht anders bei der Untersuchung von Fossilien: wir wissen aus welchen geologischen Epochen diese stammen, können evolutionäre Muster erkennen, Aussterbeereignisse ermitteln und diese Befunde mit Befunden aus anderen Teilgebieten, wie der Paläoklimaforschung, der Geophysik, Kontinentaldrift und der Untersuchung heute lebender Organismen vergleichen.
Außerdem können Paläontologen ihre Hypothesen auch mit Zahlenwerten testen. So kann man die Laufgeschwindigkeit von Dinosauriern messen, indem man fossile Fußspuren untersucht und die Schrittlänge mit der Länge der Gliedmaßen vergleicht. Man kann auch die Beißkraft der Dinosaurier ermitteln, indem man dreidimensionale digitale Modelle, basierend auf CT-Daten der Fossilien erstellt. Solche Hypothesen können dann bestätigt oder widerlegt werden. Dass aus einigen Spekulationen Realität werden konnte, zeigt die Farbgebung einiger ausgestorbener Lebewesen.
Z. B. wurde lange Zeit angenommen, dass die Farbe bei Ichthyosauriern bloße Spekulation sein konnte (Abb.9). Aber tatsächlich konnte in einigen Ichthyosaurus-Fossilien sogenannte Melanocyten in der fossilen Haut der Tiere entdeckt wurden. Melanocyten enthalten das Pigment Melanin, welches für die Farbe der Haut, Haare und Federn verantwortlich ist. Aus der Verteilung von Melanozyten bei einigen bei Holzmaden gefundenen Ichthyosauriern kann man schließen, dass die Meeresechsen dunkle Rücken und helle Bäuche hatten, um optisch mit dem Hintergrund zu verschmelzen. Außerdem zeigen die Überreste des Tieres Spuren einer Fettschicht unter der Haut, die man auch von Walen kennt und bei ihnen als Blubber bezeichnet. Diese Anpassung verhindert, dass der Körper im kalten Wasser auskühlt und belegt damit, dass die Ichthyosaurier ihre Körper aktiv erwärmten, sie also höchstwahrscheinlich gleichwarm waren (Lindgen et al., 2014; 2018).

Abb. 9: Lebendrekonstruktion eines Ichthyosaurus
Je tiefer wir in die Vergangenheit unserer Erdgeschichte gehen, desto schlechter wird der Fossilbestand. Das hat zwei Gründe. Biologisch gesehen gab es früher weniger Arten von Organismen. Geologisch gesehen sind relativ wenige Gesteine (und Fossilien) aus älteren Zeiten erhalten geblieben, weil viele durch Erhitzung, Verformung und andere Veränderungen die Fossilien zerstörten. Das frühe Leben der Erde war sicherlich mikroskopisch klein und hatte einen weichen Körper, eine sehr wenig vielversprechende Kombination für die Versteinerung.
Wenn direkte Fossilbelege fehlen, stützen sich Paläontologen auf molekularbiologische Befunde moderner Organsimen, um z. B. evolutionäre Prozesse aus der Vergangenheit besser zu verstehen.
Wie sich paläontologische und molekularbiologische Befunde gegenseitig befruchten können, soll an einem Beispiel gezeigt werden.
Die taxonomische Einordnung der Wale war lange Zeit ein Streitpunkt der Debatte bei Paläontologen. Natürlich waren sich alle Paläntologen sicher, dass Wale Säugetiere sind und von einem Vorfahren abstammen, welcher an Land lebte. Doch zu welcher Gruppe gehörten die Vorfahren der Wale?
Lange Zeit hatten Paläontologen wegen der ähnlichen Beschaffenheit von Schädel und Zähnen geglaubt, die Vorfahren der Wale seien die Mesonychia gewesen, eine Gruppe von fleischfressenden Huftieren mit umstrittener systematischer Stellung, die nicht näher mit den heutigen Huftieren verwandt sind (Abb. 10).

Abb. 10: Mesonychia
Spätere molekularbiologische Studien zeigten aber, dass Wale stammesgeschichtlich eng mit den Paarhufern (Artiodactyla) verwandt sind (Abb. 11, zur Evolution der Wale siehe Prothero 2017).
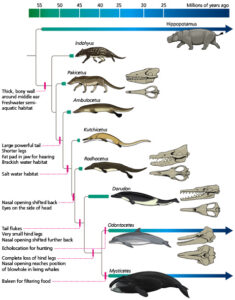
Abb. 11: Stammbaum der Wale
Diese taxonomische Einordnung wurde durch spätere Fossilfunde bestätigt. Eine Synapomorphie der Paarhufer ist das Sprungbein (Astragalus), ein Knochen im oberen Sprunggelenk (Knöchel). Der Astragalus bei Paarhufern hat eine charakteristische Form, welches durch doppelte Gelenkrollen („Doppelwalzengelenk“) gekennzeichnet ist.
Fossilfunde früher amphibisch lebender Wale haben gezeigt, dass dieses Merkmal auch bei Walen vorkommt (Abb. 12. Innerhalb der Paarhufer sind die Flusspferde molekularbiologisch die engsten Verwandten der Wale. Für diese Auffassung sprechen auch einige gemeinsame anatomische Merkmale, etwa Übereinstimmungen in der Morphologie der hinteren Backenzähne (Benton 2007: S. 360).
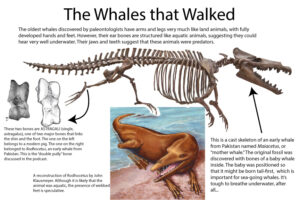
Abb. 12: Astragalus bei Urzeitwalen
Taphonomie und Aktuopaläontologie
Die Taphonomie beschäftigt sich mit der Entstehung von Fossilien und wurde vom sowjetischen Wissenschaftler Iwan Antonowitsch Jefremow im Jahr 1940 begründet. Sie greift dabei auf die Erkenntnisse verschiedener Disziplinen wie der Chemie, der Ökologie oder der Geologie zurück. Wir haben uns ja vorhin mit der Entstehung von Fossilien allgemein befasst. Die Prozesse der Einbettung, Umkristallisation und Erhaltung der Fossilien wären dementsprechend Teilbereiche der Taphonomie. Eine weitere Teildisziplin der Taphonomie ist die Aktuopaläontologie (Schäfer 1962). Diese Fachdisziplin geht davon aus, dass physikalische und biologische Prozesse in fossilen Lebens- und Ablagerungsräumen den heutigen ähnlich sind. Daher kann man durch praktische Anwendung des Aktualismus paläontologische Funde vor dem Hintergrund heute beobachtbarer Vorgänge deuten. Ziel ist es, vergangene Lebensräume und Umwelten ausgestorbener Tiere und Pflanzen anhand rezenter Umweltsituationen zu rekonstruieren.
Neben klimatischen Faktoren ist es von Bedeutung, ob die Verwesung und die Einbettung des Kadavers an freier Luft oder unter. Da die Umstände des Todes bei den einzelnen Fossilien unterschiedlich sind, ist es absolut notwendig den geologischen und paläontologischen Kontext zu analysieren. Dadurch können verwertbare Schlüsse über die damaligen Umweltverhältnisse gezogen werden (Wuttke & Reisdorf 2012). Weiterhin spielen eine Reihe von inneren und äußeren Faktoren eine Rolle, die den Kadaver bis zu seiner kompletten Einbettung beeinflussen und seine Vollständigkeit sowie den Grad seiner Erhaltung und der Verstreuung seiner Knochen bestimmen. Alle Modifikationen chemischer und physikalischer Parameter, die in einem Kadaver stattfinden, haben das Potential die Position wie auch die Exartikulation anatomischer Einheiten, also das Zerfallen in isolierte Körperteile, zu beeinflussen (Wuttke & Reisdorf 2012).
Betrachtet man viele Wirbeltierfossilien, so fallen bei ihnen die eigentümlich verkrümmten Körperhaltungen auf. So ist z. B. bei vielen Dinosaurierfossilien der Hals nach hinten gekrümmt (Abb. 13).
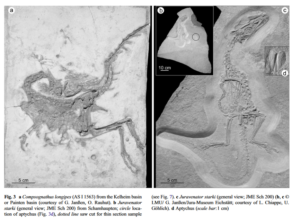
Abb. 13: gekrümmte Hälse bei Dinosaurier-Fossilien
Lange Zeit wurden die Verformungen als Folge einer Störung des zentralen Nervensystems interpretiert, die während des Todes (perimortal) auftritt. Dadurch ziehen sich die Muskeln zusammen und Hals und Rücken biegen sich krampfartig nach hinten. Diese als Opisthotonus bezeichnete Körperhaltung wurde schon 1918 von Roy Moodie (Moodie 1918) detailliert beschrieben und in einer ausführlichen Studie von Faux & Padian (2007) erneut bestätigt.
Eine 2012 erschienen Studie von Reisdorf & Wuttke (2012) stellte diese Interpretation jedoch wieder in Frage. Dabei untersuchten sie die Taphonomie von Compsognathus longipes und Juravenator starki. Beide wurden in Sedimenten eines Flachmeeres gefunden, d. h. ihre Körper sanken rasch zum Meeresgrund ab. Um den Prozess der verkrümmten Körperhaltung bei Compsognathus und Juravenator (Abb. 13) nachzuvollziehen wurde eine Hypothese von B. Dean (1919) überprüft. Diese sagt aus, dass die Krümmung des Halses durch eine aufweichende Wirbelsäule hervorgerufen wird. Dabei tauchten Reisdorf & Wuttke (2012) gerupfte Hühnerhälse in Wasser und es zeigte sich, dass sich die Hälse im Wasser nach hinten krümmten; mit zunehmender Zersetzung nahm diese Krümmung immer stärker zu. Entscheidend für die Haltung des Halses ist das Ligamentum elasticum, ein Band, das die Wirbel von Hals bis Schwanz verbindet (Abb. 14). Mit Hilfe des Ligamentum elasticum können Hals und Schwanz aufrecht gehalten werden. Es handelt sich um eine energie- und muskelkraftsparende Konstruktion. Nach dem Tod der Tiere verliert das Ligamentum seine stützende Wirkung und durch den zunehmenden Zersetzungsprozess biegen sich die Hälse weiter nach hinten. Es handelt sich hierbei also um einen postmortalen Prozess, also einem Prozess der nach dem Tod des Tieres durch Verwesung auftritt, der das Biegen der Hälse auslöst. Die Körperkonstruktion bedingt die postmortale Haltungsveränderung.
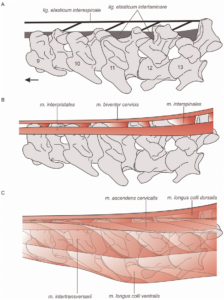
Abb. 14: in der oberen Zeichnung findet sich die Lage des Ligamentum elasticum
Wie man sieht ist die Erforschung von Fossilien nicht bloß ein Buddeln im Sand. Durch neue technische Möglichkeiten wie CT-Aufnahmen und der Molekularbiologie werden verschiedene Fachdisziplinen miteinander kombiniert und mit der Aktuopaläontologie betritt man sogar Bereiche der Forensik, die man als Otto-Normalverbraucher eher aus Sendungen wie CSI und Aktenzeichen XY ungelöst kennt.
Literatur
Benton, M. (2007): Paläontologie der Wirbeltiere
Dean B (1919) Dr. Moodies opisthotonus. Sci NS 50(1290):357
Faux CM, Padian K (2007) The opisthotonic posture of vertebrate skeletons: post-mortem contraction or death throes? Paleobiol 33:201–226
Lindgren, J. et al. (2014): Skin pigmentation provides evidence of convergent melanism in extinct marine reptiles.
Lindgren, J., Sjövall, P., Thiel, V. et al. (2018): Soft-tissue evidence for homeothermy and crypsis in a Jurassic ichthyosaur. Nature 564, 359–365. https://doi.org/10.1038/s41586-018-0775-x
Moodie RL (1918): Studies in paleopathology. III. Opisthotonus and allied phenomena among fossil vertebrates. Am Naturalist 52:384–394
Prothero, D. (1999): Fossil Record. In: Ronald Singer (Hrsg.): Encyclopedia of Paleontology. Fitzroy Dearborn Publishers, Chicago 1999, ISBN 1-884964-96-6, S. 491.
Prothero, D. (2017): Evolution – What the Fossils say and why it matters. Second edition. New York: Columbia University Press
Reisdorf, A.G., Wuttke, M. (2012): Re-evaluating Moodie‘s Opisthotonic-Posture Hypothesis in Fossil Vertebrates Part I: Reptiles – the taphonomy of the bipedal dinosaurs Comsognathus longipes and Juravenator starki from the Solnhofen Archipelago (Jurassic, Germany). Paleobio Paleoenv (2012) 92: 119 – 168
Schäfer, W. (1962): Aktuo-Paläontologie nach Studien in der Nordsee. Waldemar Kramer, Frankfurt am Main
Wuttke, M., Reisdorf, A.G. (2012): Taphonomic processes in terrestrial and marine environments. Palaeobio Palaeoenv 92, 1–3. https://doi.org/10.1007/s12549-012-0070-z
