
Intro
Hallo zusammen! Es wird mal wieder Zeit für ein neues Dinosaurier-Video. Mein letztes Video zu Dinosauriern – wenn man die sehr nerdigen Videos zur Vielfalt der Vögel einmal ausklammert – erschien im November zur Neubeschreibung von Nanotyrannus. Also definitiv zu lange her.
In diesem Video widmen wir uns erneut dem evolutionären Übergang von Nicht-Vogel-Dinosauriern zu modernen Vögeln.
Auf meinem Kanal gibt es bereits eine größere Videoreihe zur Evolution der Vögel: neun Teile mit einer durchschnittlichen Länge von je etwa 30 Minuten. Diese Reihe möchte ich nun weiter ausbauen. Ergänzt wurde sie bereits durch ein Debunking des Kreationisten Joel Tay, eine Paper-Zusammenfassung sowie ein Video zum Merkmalsmosaik von Archaeopteryx.
In Zukunft möchte ich noch etwas tiefer ins Detail gehen. Statt viele Themen in einem Video zu behandeln, konzentriere ich mich jeweils auf ein einzelnes morphologisches Merkmal und erkläre dessen evolutionäre Entwicklung.
Die Videos werden dadurch vermutlich kürzer, dafür aber zahlreicher. Einige Themen aus der ursprünglichen Reihe werde ich dabei noch einmal aufgreifen und didaktisch neu aufbereiten.
Wie viele Videos daraus am Ende werden, hängt auch ein Stück weit von eurem Interesse ab. Schreibt mir also gerne in die Kommentare, welche Themen euch besonders interessieren.
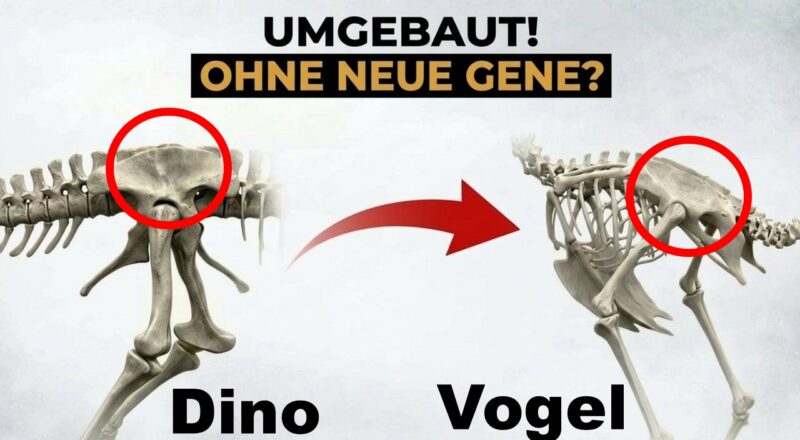
Sakralwirbel
Der Fossilbericht der Sakralwirbel
In diesem Video konzentrieren wir uns nur auf eine kleine anatomische Besonderheit, den Sakralwirbeln. Das sind die Wirbel, die sich zum Kreuzbein verschmelzen und die Wirbelsäule mit dem Becken verschmelzen.
Menschen haben übrigens 5 Sakralwirbel am Kreuzbein, Reptilien haben hingegen nur zwei.
Moderne Vögel hingegen haben zwischen 11 bis 20 Sakralwirbel. Wenn nun aber Vögel aus Dinosauriern, also Reptilien hervorgegangen sind, wie ist solch eine Änderung vonstattengegangen? Tatsächlich weichen Dinosaurier von der typischen „Reptilien haben nur zwei Sakralwirbel-Muster“ ab, denn sie haben mindestens drei. Tatsächlich ist das Vorhandensein von mindestens drei Sakralwirbel ein diagnostisches Merkmal, was, neben anderen anatomischen Merkmalen, einen Dinosaurier definiert. Damit aber nicht genug. Innerhalb einer Untergruppe der Dinosaurier, den Theropoden, also den fleischfressenden Dinosauriern, haben wir mindestens fünf bis sechs Sakralwirbel. Jetzt kommt der wirklich interessante Teil. Innerhalb der Theropoden, die mit den Vögeln am engsten verwandt sind und den frühesten Aviale, wie Archaeopteryx, Epidipteryx, Jeholornis usw. haben ebenfalls 5-6 Sakralwirbel. Bei den Vögeln mit kürzeren Schwänzen, den sogenannten Pygostyliern wie Confuciusornis und Sapeornis, sehen wir eine Zunahme der Sakralwirbel auf 7, bei den etwas weiter entwickelten Ornithoraces sehen wir 8 und schließlich bei den Ornithurae 10-11. Moderne Vögel haben hingegen 11 oder mehr Sakralwirbel.
Man kann fast jedes anatomische Merkmal auswählen, es anhand der Fossilienfunde verfolgen und beobachten, wie es sich vom Zustand der Nicht-Vogel-Dinosaurier zu dem Zustand entwickelt, den wir bei modernen Vögeln sehen, mit mehreren Zwischenstadien dazwischen.
Diese Belege stimmen vollkommen mit Evolution überein. Wir sehen, wie die Dinosaurier der Anzahl der Sakralwirbel erhöhen, dann sehen wir, wie die frühesten Vögel sich in ihrer Sakralanzahl mit den Dinosauriern überschneiden, und dann sehen wir eine allmähliche Zunahme, bis wir bei 11 angelangt sind.
Wie werden Wirbel in der Embryonalentwicklung gebildet?
Was bringt es allerdings, solche Veränderungen in Fossilien nachzuweisen, wenn wir nicht erklären können, wie sie durch Mutationen entstehen? Genau darum geht es in diesem Beitrag.
Wie können Mutationen größere Veränderungen im Körperbau hervorrufen? Häufig geschieht das über Veränderungen in regulatorischen Genen. Diese Gene steuern die Embryonalentwicklung und bestimmen, wann, wo, wie lange und wie stark andere Gene aktiv sind. Um die Zahl der Sakralwirbel zu erhöhen, braucht es weder neue Knochen noch neue Gene. Stattdessen können benachbarte Wirbel ihre Identität verändern. Wirbel ober- oder unterhalb des Kreuzbeins – etwa Lenden- oder Schwanzwirbel – werden in das Kreuzbein integriert und entwickeln entsprechende Merkmale.
Um zu verstehen, wie das möglich ist, müssen wir uns zunächst ansehen, wie Wirbel im Embryo überhaupt angelegt werden.
Die embryonalen Vorläufer der Wirbel nennt man Somiten. Das sind segmentierte Zellblöcke entlang der Rückenseite eines Wirbeltierembryos. Aus ihnen entstehen später die Wirbelkörper sowie zugehörige Gewebe wie Rippen und Muskeln.
Die meisten Wirbeltiere besitzen fünf Wirbeltypen: Hals-, Brust-, Lenden-, Sakral- und Schwanzwirbel. Welche dieser Wirbelarten aus einem Somiten entsteht, wird durch bestimmte Kombinationen sogenannter Hox-Gene festgelegt. Hox-Gene sind eine stark konservierte Familie regulatorischer Gene. Sie fungieren als eine Art Master-Regulatoren der Embryonalentwicklung und bestimmen den grundlegenden Bauplan des Körpers. Dabei steuern sie, welche Körpersegmente entlang der Längsachse entstehen und welche Identität sie annehmen.
Bemerkenswert ist, dass die Reihenfolge der Hox-Gene auf dem Chromosom der Reihenfolge ihrer Aktivierung im Embryo entspricht. Vereinfacht gesagt: vordere Hox-Gene steuern vordere Körperabschnitte, hintere Hox-Gene die hinteren.
Alle Tiere besitzen Hox-Gene. Wirbellose Tiere wie die Fruchtfliege haben jedoch nur einen Hox-Gen-Cluster mit acht Genen, während Wirbeltiere vier Cluster mit insgesamt 39 Genen besitzen – ein Ergebnis früherer Genduplikationen. Auf meinem Kanal gibt es bereits mehrere Videos zu Hox-Genen und Embryonalentwicklung, falls ihr tiefer einsteigen möchtet.
Die Identität eines Somiten – und damit eines Wirbels – wird durch eine bestimmte Kombination aktiver Hox-Gene festgelegt. Schon kleine regulatorische Veränderungen können diese Grenzen um ein oder zwei Segmente verschieben. Dadurch kann sich beispielsweise ein Lenden- oder Schwanzwirbel zu einem Sakralwirbel entwickeln.
Doch was aktiviert eigentlich ein Hox-Gen?
Die entscheidenden Signale liefern sogenannte Morphogene. Sie bilden im Embryo chemische Konzentrationsgradienten. Je nachdem, wie hoch die Konzentration eines Morphogens in einer Zelle ist, werden bestimmte Gene aktiviert oder unterdrückt. Viele dieser Morphogene sind schon im Zellplasma der Eizelle enthalten und werden nach der Befruchtung durch ein Spermium aktiviert.
Zellen „lesen“ also gewissermaßen ihre Position im Embryo anhand der Morphogenkonzentration. Hohe und niedrige Konzentrationen führen zu unterschiedlichen Genaktivitäten und damit zu unterschiedlichen Zellschicksalen. Auf diese Weise entstehen klare Grenzen zwischen Körpersegmenten, in denen jeweils andere Gene aktiv sind. Zu den wichtigsten Morphogenen in diesem Prozess gehören Retinsäure (RA), Wnt und FGF.
Am Anfang der Entwicklung besteht der Embryo aus vielen identischen, noch undifferenzierten Stammzellen. Sie befinden sich in einer Art chemischer Umgebung aus verschiedenen Morphogenen. Die Zusammensetzung dieser Umgebung bestimmt, welche Gene in einer Zelle aktiviert werden und zu welchem Zelltyp sie sich entwickelt.
Mit Beginn der Gastrulation, wenn sich der sogenannte Primitivstreifen bildet und die Zellen beginnen, sich zu spezialisieren, entsteht eine erste räumliche Ordnung im Embryo:
Im vorderen Bereich sammeln sich Retinsäure-Signale, im hinteren Bereich dominieren Wnt- und FGF-Signale. Dadurch entsteht erstmals eine klare Vorne-Hinten-Achse.
Gene, die durch Wnt aktiviert werden, steuern vor allem hintere Körperstrukturen wie Hüften oder Schwanz. Gene, die durch Retinsäure aktiviert werden, steuern dagegen vordere Strukturen wie Gehirn und Augen. FGF wiederum reguliert vor allem, wann Zellen in die nächste Entwicklungsphase übergehen.
Hox-Gene reagieren nun auf diese Morphogensignale. Jedes Hox-Gen wird nur bei bestimmten Morphogenarten und Konzentrationsschwellen aktiviert. Beispielsweise kann eine steigende Retinsäurekonzentration nacheinander verschiedene Hox-Gene aktivieren:
Wenn z. B. die Konzentration von Retinsäure über einen Wert von X steigt, wird das Gen HoxA5 aktiviert, bei einem Schwellenwert von Y HoxA7 und bei einem Schwellenwert von Z HoxA10. Jedes dieser Gene reagiert also auf einen leicht anderen Konzentrationsbereich.
Entlang der Körperachse bedeutet das: Hohe Retinsäurekonzentrationen aktivieren eher vordere Hox-Gene, während bei niedrigeren Konzentrationen zunehmend weiter hinten liegende Hox-Gene aktiv werden. In umgekehrter Richtung übernehmen Wnt-Signale diese Rolle für die hinteren Bereiche des Embryos.
Wichtig ist: Hox-Gene bauen keine Strukturen direkt auf. Sie wirken als übergeordnete Regulatoren. Dazu aktivieren sie Transkriptionsfaktoren, die wiederum andere Gene ein- oder ausschalten. Diese nachgeschalteten Gene steuern dann die Ausbildung konkreter Strukturen im Körper.
Da unterschiedliche Hox-Gene jeweils andere Zielgene aktivieren, entsprechen sie verschiedenen Abschnitten des Körpers.
Die Hox6-Gruppe steuert z. B. die Identität der Brustwirbel, unter anderem indem sie die Bildung von Rippen auslöst. Die Hox10-Gruppe unterdrückt dagegen die Rippenbildung und bestimmt damit die Lendenwirbel. Die Hox11-Gruppe legt schließlich die Identität der Sakralwirbel fest.
Wenn sich nun der Aktivitätsgradient eines Hox-Gens leicht verschiebt, verschiebt sich auch die Grenze, an der diese Gene aktiv sind. Dadurch kann ein Wirbel, der ursprünglich ein Lendenwirbel geworden wäre, stattdessen die Eigenschaften eines Sakralwirbels annehmen.
Eine solche Verschiebung kann entstehen, wenn sich zum Beispiel der Wnt-Morphogengradient leicht verändert oder wenn ein Hox-Gen bereits bei etwas geringerer Wnt-Konzentration aktiviert wird. In beiden Fällen erweitert sich sein Expressionsbereich.
Die Hox11-Gene aktivieren Programme, die Wirbeln typische Sakralmerkmale verleihen – etwa große Querfortsätze und das Iliosakralgelenk, also die knöcherne Verbindung zwischen Wirbelsäule und Becken.
Um bei Vögeln mehr Sakralwirbel zu erhalten, musste daher nichts Neues entstehen. Es reichte, dass sich die Expression von Hox11 etwas weiter nach vorne ausdehnte. Dadurch fielen zusätzliche Wirbel in diesen Expressionsbereich und erhielten die Identität eines Sakralwirbels.
Mit anderen Worten: Es mussten keine neuen Gene, keine neuen Knochen und keine zusätzlichen Somiten entstehen. Bereits vorhandene Wirbel veränderten einfach ihre Identität – durch kleine Änderungen in der Genregulation während der Embryonalentwicklung.
Wie kann eine neue Wirbelidentität entstehen?
Erstens: Die Gene, die Wnt-Morphogene produzieren, könnten stärker exprimiert werden. Dadurch würde sich der Wnt-Konzentrationsgradient weiter ausdehnen und Gene wie Hox11, die durch Wnt aktiviert werden, könnten in einem größeren Bereich aktiv sein. Umgekehrt könnte auch die Expression von Morphogenen im vorderen Embryobereich abnehmen, die hintere Programme normalerweise unterdrücken. Auch das würde die Aktivität hinterer Hox-Gene erweitern.
Zweitens: Veränderungen an Zellrezeptoren. Wenn Gene, die Wnt-Rezeptoren codieren, stärker exprimiert werden oder empfindlichere Rezeptoren entstehen, können Zellen mehr Wnt-Signale wahrnehmen. Dadurch können auch Bereiche mit eigentlich niedriger Wnt-Konzentration ausreichend Signal erhalten, um Hox11 zu aktivieren. Das Ergebnis wäre ebenfalls ein größerer Expressionsbereich.
Drittens: Veränderungen in regulatorischen DNA-Sequenzen, etwa durch die Duplikation von Enhancern. Enhancer sind kurze DNA-Abschnitte, an die Transkriptionsfaktoren binden und so die Aktivität eines Gens verstärken. Wenn sich solche Enhancer bei der DNA-Replikation verdoppeln, können mehr Transkriptionsfaktoren binden und das entsprechende Gen wird häufiger oder stärker aktiviert.
Enhancer-Duplikationen sind ein bekannter evolutionärer Mechanismus, um Genexpression zu verändern, besonders bei Entwicklungsgenen.
Wenn dadurch zum Beispiel Hox11 stärker oder in einem größeren Bereich aktiv ist, können zusätzliche Wirbel die Eigenschaften von Sakralwirbeln annehmen.
Das kann sogar einen funktionellen Vorteil haben. Eine größere Zahl von Sakralwirbeln erhöht die Stabilität des Rumpfes und die Kraftübertragung zwischen Wirbelsäule und Becken. Gerade bei Vögeln kann das die Effizienz von Bewegung und Flug verbessern.
Individuen mit solchen Veränderungen können daher erfolgreicher Nahrung finden, schneller fliehen und sich häufiger fortpflanzen. Über viele Generationen hinweg kann sich eine solche Mutation so im Genpool der Population durchsetzen.
Laborversuche mit Mäusen und Hühnern zeigen, dass sich Wirbeltypen tatsächlich verändern lassen, wenn man die Expressionsbereiche von Hox-Genen experimentell verschiebt. Durch gezielte genetische Veränderungen konnten Forscher Wirbel in andere Wirbeltypen umwandeln. Es handelt sich also nicht nur um eine theoretische Erklärung – der zugrunde liegende Mechanismus wurde im Labor bestätigt.
Sowohl Dinosaurier als auch Vögel haben ihre Zahl an Sakralwirbeln vermutlich schrittweise erhöht, indem benachbarte Wirbel in das Kreuzbein integriert wurden. Möglich wurde das durch eine Ausdehnung des Hox11-Expressionsbereichs.
Dieses Beispiel zeigt ein allgemeines Prinzip der Evolution: Große Veränderungen im Körperbau können durch kleine Anpassungen in der Embryonalentwicklung entstehen. Schon eine etwas frühere Aktivierung eines Gens, eine längere Aktivitätsdauer oder eine leicht veränderte Expressionsstärke kann den Phänotyp deutlich verändern.
Dafür braucht es meist keine neuen Gene, sondern nur kleine Veränderungen in den regulatorischen Sequenzen, die steuern, wann und wo Gene aktiv sind.
Vielen Dank, dass ihr mich auf dieser Reise begleitet habt.
Bis zum nächsten Mal!
Welches Merkmal der Dinosaurier soll ich als Nächstes behandeln? Ich habe schon einige Ideen, aber schreibt mir eure Vorschläge gerne in die Kommentare. Und wenn euch das Video gefallen hat, lasst gern ein Like und ein Abo da – mehr Dino-Content folgt garantiert.
Literatur
Literatur zu den Merkmalen der Dinosaurier:
Cau, A. (2018): The assembly of the avian body plan: a 160-million-year long process. Bollettino della Società Paleontologica Italiana. 57 (1): 1–25. https://www.researchgate.net/publication/324941372_The_assembly_of_the_avian_body_plan_A_160-million-year_long_process
Fastovsky, D. E., Weishampel, D. B. (2021): Dinosaurs: A Concise Natural History (4th ed.). Cambridge university Press, Cambridge.
Schweitzer, M. H., Schroeter, E. R., Czajka, C. D. (2021): Dinosaurs How We KNow What We Know. CRC Press
Literatur zu Hox-Genen, Entstehung der Wirbelsäule aus Somiten und Evo-Devo
Als Überblick mein Video: Wie Embryonen unsere Evolution steuern (Von LUCA bis Eva #6) https://www.youtube.com/watch?v=B8Osnw6ioRU
Afzal, Z, Krumlauf, R (2022). Transcriptional Regulation and Implications for Controlling Hox Gene Expression. J. Dev. Biol. 10, 4. https://doi.org/10.3390/jdb10010004
Boekhout, M. Posterior development, a growing body of evidence. https://www.researchgate.net/publication/41104998_Posterior_development_a_growing_body_of_evidence
Mallo, M. (2016), Revisiting the involvement of signaling gradients in somitogenesis. FEBS J, 283: 1430-1437. https://doi.org/10.1111/febs.13622
Wellik DM (2007). Hox patterning of the vertebrate axial skeleton. Dev Dyn.236(9):2454-63. https://pubmed.ncbi.nlm.nih.gov/17685480/
