

In dieser Episode eröffnet sich uns ein bemerkenswert vielfältiges Spektrum an Vogelgruppen, die auf den ersten Blick nur wenig gemeinsam zu haben scheinen, deren gemeinsame Wurzeln jedoch tief in der Evolution verwurzelt sind. Zu diesen Gruppen gehören die Nashornvögel, die Trogone, Kurole, Mausvögel und Wiedehopfe. Sie werden zusammen mit den Spechtvögeln, Eisvögeln und Racken als Coraciimorphae zusammenfasst.
Mausvögel (Coliidae)

Mausvögel sind die einzige Familie innerhalb ihrer eigenen Ordnung und kommen mit nur sechs Arten in zwei Gattungen ausschließlich im tropischen Afrika vor, vornehmlich in den Savannen (Winkler et al. 2015, 2020a, Del Hoyo 2020, Del Hoyo et al. 2001). Ihren Namen verdanken sie, analog zu den Mäusen, ihrer geselligen Lebensweise und der Bewegungsart, wie eine Maus durch das Gebüsch zu huschen. Sie weisen außerdem ein fast fellartiges wirkendes graues bis bräunliches Gefieder auf. Sie fressen überwiegend Beeren und Früchte und kommen nur ausnahmsweise auf den Boden. Sie werden je nach Art 29 – 38 cm lang, aber die Hälfte der Körperlange macht der Schwanz aus, wodurch die Tiere insgesamt doch recht klein sind. Ähnlich wie die Kolibris können Mausvögel in einen nächtlichen Starreschlaf, also Tropor, fallen. In diesem energiesparenden Schlaf sind Körpertemperatur und Stoffwechsel stark reduziert.
Trotz ihrer geringen Artenzahl und beschränkten Verbreitung, sind die heutigen Arten nur ein Bruchteil der einstigen Vielfalt dieser Vogelgruppe, die einst in Nordamerika und Europa weit verbreitet war. Fossil sind 16 Gattungen bekannt, die neben einigen basalen Formen in vier Familien aufgeteilt werden. Neben den eigentlichen Mausvögeln zählen hierzu die Sandcoleidae, Selmeidae und die Chascacocoliidae (Mayr 2000, 2001, 2005, 2009, 2010, 2013, 2015, 2016, 2018, Mayr & Mourer-Chauviré 2004, Mayr & Peters 1998, Houde & Olson 1992, Ksepka & Clarke 2009, 2010, Ksepka et al. 2017, Peters 1999, Saupe et al. 2019, Zelenkov & Dyke 2008).
Die ursprünglichsten Sandcoleidae stammen aus dem Paläozän und Eozän Nordamerikas und Europas (Sandcoleus, Anneavis, Eoglaucidium). Sie unterscheiden sich von den Mausvögeln z. B. durch längere Oberarmknochen und einen kurzen Tarsometatarsus mit einem spezialisierten Greiffuß. Die Selmeidae sind mit nur einer Art Selmes absurdipes aus dem Eozän Europas bekannt, der in seiner Skelettmorphologie den heutigen Mausvögeln insgesamt ähnelt, aber einen längeren Schnabel und kürzere Zehenknochen hat. Die Familie der Chasacocoliidae hat zwei Arten in einer Gattung aus dem Eozän von Nordamerika und Europa (Chascacocolius oscitans und Chascacocolius cacicirostris). Sie haben einen sehr langen, hinter dem Ohr befindlichen, Fortsatz des Unterkiefers, welcher die Hebelwirkung der Kiefermuskeln verstärkt. Diese Fortsätze sind bei modernen Mausvögeln nicht mehr vorhanden, finden sich aber in einigen ausgestorbenen Mitgliedern der Familie, wie Masillacolius aus dem Eozän Deutschlands. Sie wurden also bei den modernen Mausvögeln reduziert. Celericolius aus der eozänen Green-River-Formation Nordamerikas ähnelt modernen Mausvögeln in der allgemeinen Skelettmorphologie, hat aber proportional längere Flügel.
Weiter fortgeschrittene Mausvögel gehören zu einer Gruppe, zu der auch die heute lebenden Arten gehören und die durch einen großen Fortsatz des Carpometacarpus gekennzeichnet sind. Die ältesten Vertreter dieser moderneren Stammgruppe gehören zu Primocolius und Paleaospiza aus dem Eozän Frankreichs bzw. Nordamerikas. Ein weiteres basales Mausvogel-Taxon, Oligocolius, existierte jedoch noch im Oligozän in Deutschland. Seine Flügel waren länger als die modernen Mausvögel und sein Schnabel war Papagei-ähnlich. Es finden sich auch große Samen im Bereich der Speiseröhre, die auf einen Kropf hindeuten, der bei modernen Mausvögeln fehlt.
Das Verschwinden der Mausvögel aus Europa ist wahrscheinlich auf die klimatische Abkühlung gegen Ende des Miozäns zurückzuführen, da die aufkommenden kalten Wintermonate das Überleben von nicht wandernden, fruchtfressenden Vögeln in nördlichen Breiten nicht mehr zuließen. In Afrika, wo heute nur noch Mausvögel vorkommen, stammt der früheste fossile Nachweis aus dem frühen Miozän von Namibia.
Kurole (Leptosomidae)
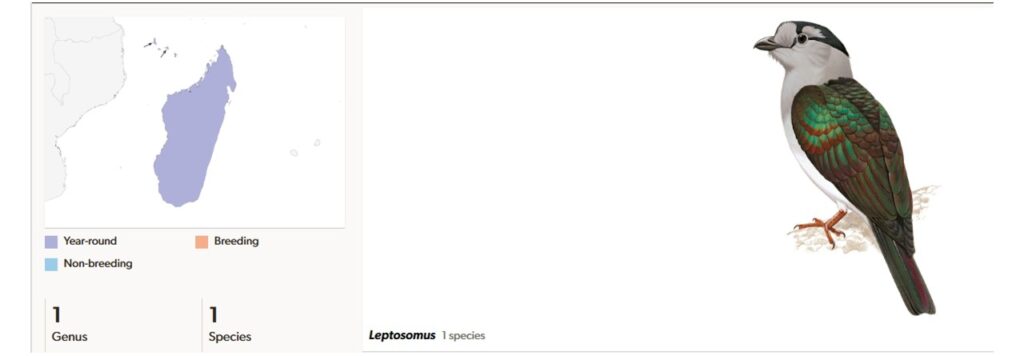
Eine weitere Ordnung von Afroaves, die in prähistorischer Zeit wesentlich weiter verbreitet waren ist die Ordnung der Leptosomiformes, bestehend aus nur einer Familie mit einer Art: Leptosomus discolor, der Kuckucksroller oder auch Kurol genannt. Er kommt nur auf Madagaskar und den Komoren vor (Winkler et al. 2020b, 2015, Del Hoyo et al. 2001, Del Hoyo 2020).
Sie werden 38 bis 50 Zentimeter groß und erreichen ein Gewicht von 192 bis 270 Gramm. Im Aussehen besteht zwischen Weibchen und Männchen ein ausgeprägter Geschlechtsdimorphismus. Das Weibchen ist größer als das Männchen und hat ein rotbraunes bis bräunliches Oberseitengefieder; an der Unterseite ist es gelbbraun. Das Rückengefieder ist schwarz gebändert, während die Unterseite schwarz gefleckt ist. Beim Männchen ist das Rückengefieder grün mit einem kupferfarbenen Metallglanz. Der Nacken und der Kopf sind grau und das restliche Federkleid hat eine hellblaue über hellgraue bis graue Farbe. Die Schnäbel beider Geschlechter sind grauschwarz bis schwarz gefärbt. Die Beine sind im Verhältnis zum Körper recht kurz. Kurole sind gute Flieger und ihre Nahrung besteht vorwiegend aus Insekten und kleinen Reptilien. Unverkennbar sind auch die Rufe der Vögel, die zu den typischen Vogelgesängen der Wälder Madagaskars gehören.
Fossile Stammgruppen der Ordnung Leptosomiformes sind aus dem Eozän Nordamerikas und Europas bekannt und werden der Gattung Plesiocathartes mit fünf Arten zugeordnet, welche dem Kurol recht ähnlich sind. Diese Stammgruppe belegt zum einen die viel weitere Verbreitung dieser Vogelordnung und dazu noch eine größere Vielfalt (Mayr 2009, 2016). Möglicherweise starben aber diese Gattungen schon im Oligozän aus, sodass diese Ordnung nur noch auf Madagaskar und den Komoren vorkommt.
Trogone (Trogonidae)
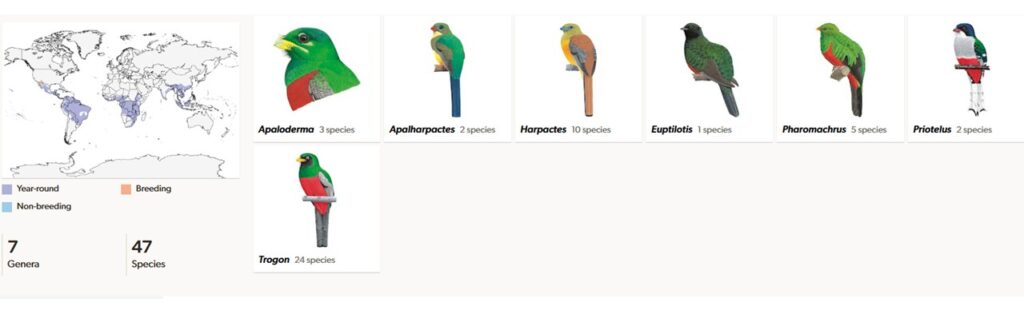
Die Ordnung der Trogoniformes besteht zwar, wie die vorher genannten, auch aus nur einer Familie (Winkler et al. 2020c, 2015, Del Hoyo et al. 2001, Del Hoyo 2020). Doch mit knapp 50 Arten in sieben Gattungen und drei Unterfamilien sind sie deutlich vielfältiger als die Mausvögel und Kurole. Ihre Verbreitung beschränkt sich auf die tropischen Gebiete von Mittel- und Südamerika, Afrika und Asien. Die Unterfamilie Aplodermatinae hat drei Arten in einer Gattung, die in Afrika vorkommen. Die Unterfamilie Harpactinae hat 2 Gattungen mit insgesamt 12 Arten, die im tropischen Asien vorkommen und die Unterfamilie Trogoninae kommt mit über 30 Arten in vier Gattungen in Mittel- und Südamerika vor, wobei das Verbreitungsgebiet des Kieferntrogons (Euptilotis neoxenus) und des Kupfertrogons (Trogon elegans) bis nach Arizona und New Mexiko in den USA reicht.
Trogone gehören zu den farbenprächtigsten Vögeln. Vor allem die Männchen zeigen sich in einem Prachtkleid, die Weibchen sind weniger auffällig gefärbt. In der dünnen, leicht reißenden Haut sitzen die Federn sehr locker. Das Gefieder hat auf der Oberseite meist eine leuchtende, metallisch schimmernde, smaragdgrüne oder blaue Farbe. Auf der Unterseite ist das Gefieder oft rot, blau, gelb oder weiß. Sie besitzen abgerundete Flügel und lange Schwanzfedern. Trogone haben heterodactyle Füße: die erste und zweite Zehe weisen nach hinten. Bei anderen Baumvögeln weisen hingegen die erste und vierte Zehe nach hinten, das wären sog. zygodactyle Füße. Sie ernähren sich von Insekten, anderen Wirbellosen, Beeren und Früchten. An ihren kurzen, breiten und kräftigen Schnäbeln befinden sich auf jeder Seite der Schnabelwurzel steife Borsten, die eine Art Fanggerät für fliegende Beutetiere bilden. Trogone sind Höhlenbrüter. Mit dem Schnabel und den Füßen wird in einem morschen Baum oder in einem Termitenhügel eine Höhle gegraben oder sie nehmen eine verlassene Nisthöhle ein. Das Gelege, das aus zwei bis vier Eiern besteht, wird von beiden Altvögeln 15 bis 19 Tage lang ausgebrütet. Die Jungvögel verlassen das Nest nach 16 bis 30 Tagen. Die meisten Trogonarten leben in tropischen Regenwäldern, doch auch hier gibt es Ausnahmen. Der afrikanische Narnia-Trogon (Apaloderma narina) bewohnt außer Regenwäldern auch Trockenwälder und Savannen. Die mittelamerikanischen Kupfertrogon (Trogon elegans) und Kieferntrogon (Euptilotis neoxenus) kommen zwar vorwiegend in Wäldern vor, erreichen aber auch die temperierten Zonen von Arizona und New Mexiko in den USA, wobei der Kieferntrogon auch trockenere Habitate bewohnen kann. Die bekannteste Trogon-Art dürfte der in den Gebirgsregenwäldern von Mittelamerika lebende Quetzal (Pharomachrus mocinno) sein, der auch der Nationalvogel von Guatemala ist. Der Quetzal wurde von den präkolumbischen Kulturen wegen seiner prachtvollen langen Schwanzfedern, die als Kopfschmuck für ihre Fürsten und Priester dienten, gejagt. Beispielhaft ist die Verwendung seiner Federn bei der sogenannten Federkrone Moctezumas. Für die Azteken in Mexiko war der Vogel eng mit der Gottheit Quetzalcoatl verbunden und wurde daher nicht getötet. Wer einen Quetzal tötete, wurde mit dem Tode bestraft.
Fossil waren Trogone weiterverbreitet als heute. Der älteste Nachweis ist Septentrogon aus dem Eozän. Aus dem Oligozän Frankreichs stammt Primotrogon, während Masillatrogon aus der Grube Messel (Eozän) bei Darmstadt stammt. Beim 2023 beschriebenen Eotrogon stenorhynchus, aus dem frühen Eozän Englands konnte schon die für die Familie typischen heterodaktylen Füße nachgewiesen werden (Kristoffersen 2002, Mayr 1999, 2009, 2009a, 2016, 2023).
Die heutigen Trogone können sich nur schlecht über große Entfernungen ausbreiten, und nach den ähnlichen Flügelskeletten zu urteilen, scheint dies auch für ihre ausgestorbenen Verwandten zu gelten. Das Verschwinden der Trogone aus den nördlichen Breitengraden ist daher, wie bei den Mausvögeln und Kurolen, auf das kühlere Klima und größere Saisonalität ab dem Miozäne zurückzuführen.
Ihre Fähigkeit sich schlecht auszubreiten erklärt, warum es keine Trogone in abgelegenen Inseln wie Madagaskar und der australischen Region gibt. Madagaskar bereits zu weit von einem Kontinent entfernt, als dass sich Trogone hätten ausbreiten können. Das Fehlen in der australischen Region könnte mit der Wallace-Linie zusammenhängen, eine geographische Barriere in Indonesien, die auch andere Faunengruppen voneinander trennt.
Molekulare Daten belegen, dass die afrikanische Unterfamilie Apalpderma die ursprünglichste und zugleich Schwestergruppe der beiden anderen Unterfamilien ist (Johansson & Ericson 2005, Sangster et al. 2023). Dies spricht für einen Ursprung in der Alten Welt. Wann sich die Trogone nach Amerika ausgebreitet haben, ist nicht bekannt, aber die Tatsache, dass alle paläogenen Trogonfossilien zu Vertretern der Stammgruppe gehören, deutet darauf hin, dass die Ausbreitung nicht vor dem Neogen erfolgte.
Wiedehopfe (Upupidae)

Die Ordnung der Bucerotiformes hat drei oder vier rezente Familien: die Wiedehopfe, Baumhopfe und die Nashornvögel. Baumhopfe und Hopfe werden oft als Upupiformes zusammengefasst. Von einigen Autoren werden die Hornraben als eigene Familie außerhalb der Nashornvögel angesehen. Dazu kommen noch zwei ausgestorbene Familien.
Wiedehopfe sind eine artenarme Familie, die nach aktuellem Stand drei einander recht ähnliche Arten umfassen (Winkler et al. 2020d, 2015, Del Hoyo et al. 2001, Del Hoyo 2020). Der Eurasische Wiedehopf (Upupa epops), der in Europa, fast ganz Asien und in Afrika nördlich des Äquators vorkommt, der Afrikanische Wiedehopf (Upupa africana), der in Afrika südlich des Äquators vorkommt und der auf Madagaskar heimische Madagaskar-Wiedehopf (Upupa marginata). Früher wurden diese drei Arten als eine Art zusammengefasst. Eine weitere anerkannte Art, der St.-Helena-Wiedehopf (Upupa antaios), starb im 16. Jahrhundert aus (Olson 1975). Wiedehopfe sind unverkennbare Vögel mit großer, aufrichtbarer Federhaube. Ihre Flügel sind breit und rundlich, der Schwanz rechteckig. Flügel und Schwanz sind auffällig schwarz-weiß gezeichnet, der Körper ist variabel rosabraun gefärbt. Ihr langer, dünner Schnabel ist nach unten gebogen. Der bevorzugte Lebensraum ist offenes baumbestandenes Gelände, wie etwa Savanne oder Kulturland. Sie ernähren sich überwiegend von Insekten und anderen Wirbellosen. Oberflächlich sehen sich die verschiedenen Arten recht ähnlich, unterscheiden sich aber in der Federfärbung und -musterung. Außerdem gibt es Unterschiede im Gesang und in der Größe der Vögel.
Beim plötzlichen Auftauchen eines Greifvogels nehmen Wiedehopfe eine Tarnstellung ein, in die sie mit breit gespreizten Flügeln und Schwanz flach auf dem Boden liegen; Hals, Kopf und Schnabel sind steil nach oben gerichtet. Ihr kontrastreiches Gefieder verschmilzt so mit der Umgebung. Küken im Nest sind dafür bekannt wie Schlangen zu zischen oder Kot aus ihrer Nesthöhle zu spritzen. Besonders wirkungsvoll scheint jedoch das Absondern eines sehr übel riechenden Sekretes aus der Bürzeldrüse zu sein (Martín-Vivaldi et al. 2010, Martín-Platero et al. 2006). Der eurasische Wiedehopf ist in den gemäßigten Zonen ein Zugvogel und überwintert in Afrika oder Südasien. Die Bestände des eurasischen Wiedehopfes sind in vielen Ländern durch Biotopzerstörung und Pestizideintrag gefährdet. Aus diesem Grund ist die Art in vielen Ländern gesetzlich geschützt.
Baumhopfe (Phoeniculidae)

Nah verwandt mit den Wiedehopfen sind die Baumhopfe, die in 8 oder 9 Arten in zwei Gattungen nur in Afrika südlich der Sahara vorkommen (Winkler et al. 2020e, 2015, Del Hoyo et al. 2001, Del Hoyo 2020). Je nach Art sind sie zwischen 24 und 40 cm lang, wobei die Hälfte der Körperlänge ihr langer Schnabel und der Schwanz ausmachen. Im Gegensatz zu den Wiedehopfen besitzen sie ein metallisch glänzendes Gefieder, oft schwarz, blau, grün oder violett, und es fehlt ihnen die aufrichtbare Haube. Auf den Flügeln und auf dem Schwanz finden sich weiße Flecken. Baumhopfe haben kurze Beine, ihre Füße tragen Krallen, die viel länger sind als die der Wiedehopfe. Die Geschlechter unterscheiden sich kaum. Die meisten Arten bewohnen die Savannen des Kontinents, wobei zwei Arten: Waldhopf (Phoeniculus castaneiceps) und Weißmaskenhopf (Phoeniculus bollei) die Regenwälder bewohnen. Auf der Futtersuche bewegen sie sich ruckartig an Stämmen und Ästen aufwärts und lassen sich dann wieder nach unten fallen oder fliegen zum nächsten Baum. Beim Klettern stützen sie sich auf ihren Schwanz, dessen Federn gestuft, aber nicht versteift sind. Sie fressen vor allem Insekten und deren Larven und Eier, seltener Knospen und Samen. Sie sind Höhlenbrüter und legen zwei bis fünf grünliche oder bläuliche Eier, die 16 bis 18 Tage allein vom Weibchen bebrütet werden. Während der Brutzeit bildet die Bürzeldrüse der Baumhopfe, den Wiedehopfen ähnlich, eine moschusartig riechende Flüssigkeit.
Messelirrisoridae & Laurillardiidae
Fossile Bucerotiformes stammen überwiegend aus der Alten Welt, aber es gibt ein unbeschriebenes Skelett aus der Green River Formation, das möglicherweise einen Nachweis dieser Vögel aus der Neuen Welt darstellt (Mayr 2016). Die ältesten gut untersuchten Fossilien gehören zu den Messelirrisoridae, die im Eozän in Europa vorkamen und besonders häufig in der Grube Messel zu finden sind (Mayr 2009, 2016). Es sind kleine Vögel und einige Arten erreichten nur die Größe eines durchschnittlichen modernen Kolibris. Sie weisen eine Reihe von abgeleiteten Merkmalen der Wiedehopfe und Baumhopfe auf, doch fehlen ihnen auch einige der Apomorphien, die die Bucerotiformes gemeinsam haben (Mayr 1998, 2006a). Sie weisen zudem Unterschiede in der Fußmorphologie auf, so einen kurzen Tarsometatarsus und eine sehr lange Hinterzehe. Man geht davon aus, dass sie eher auf Bäumen sitzend verbracht haben und keine spezialisierten Stammkletterer wie Baumhopfe oder Bodenjäger wie Wiedehopfe waren.
Eine weitere Stammgruppenfamilie sind die Laurillardiidae aus dem Oligozän Europas, die mit den Wiedehopfen und Baumhopfen näher verwandt sind als die Messelirrisoridae (Mayr et al. 2019). Hierbei handelt es sich um recht kleine Vögel. Weiterhin ist die kleinwüchsige Gattung Phirriculus aus dem Miozän Frankreichs und Deutschlands bekannt (Mlíkovský & Göhlich 2000), die möglicherweise ein Vertreter der Baumhopfe ist. Diese kleinwüchsigen Arten legen nahe, dass sie über Millionen Jahre hinweg in Europa mit den Sperlingsvögeln koexistierten und stellen frühere Hypothesen in Frage, wonach die Ankunft der Sperlingsvögel in Europa zum Aussterben der kleinen, nicht sperlingsförmigen Baumvögel führte. Es ist aber nach wie vor rätselhaft, warum kleine Baumhopf/Wiedehopf-Verwandte in der nördlichen Hemisphäre ausgestorben sind. Auch wenn diese insektenfressenden Vögel nach dem Einsetzen der kalten Winter in der nördlichen Hemisphäre möglicherweise nicht mehr genügend Nahrung fanden, bleibt die Frage offen, warum sie nicht in den tropischen Regionen Asiens vorkommen, sondern heute auf Afrika südlich der Sahara beschränkt sind.
Nashornvögel und Hornraben (Bucerotidae und Bucorvidae)

Die bekanntesten und auffälligsten Familien der Bucerotiformes sind die Nashornvögel. Aktuell werden sie in zwei Familien aufgeteilt: die afrikanischen Hornraben die in Afrika und Asien vorkommenden Echten Nashornvögel (Winkler et al. 2020f, 2015, Del Hoyo et al. 2001, Del Hoyo 2020).
Hornraben, lange Zeit als Gattung (Bucorvus) und Unterfamilie innerhalb der Nashornvögel angesehen kommen in zwei Arten – Nördlicher (Bucorvus abyssinicus) und Südlicher Hornrabe (Bucorvus leadbeateri) – in den Savannen Afrikas vor und sind hauptsächlich bodenbewohnende Vögel. Anders als die nahe verwandten Nashornvögel sind Hornraben gute Läufer und können sich mit bis zu 30 km/h fortbewegen. Hornraben haben 15 Halswirbel, während die Nashornvögel lediglich 14 aufweisen. Die Beine sind außerdem länger und sie weisen eine spezielle Sehne auf, die vom Becken zum Oberschenkelknochen verläuft. Die Tiere erreichen eine Flügelspannweite von fast zwei Metern und ein Gewicht von durchschnittlich vier Kilo. Unterscheidungsmerkmal bei beiden Arten ist vor allem die Färbung der unbefiederten Gesichtspartien, die beim Nördlichen Hornraben blau und beim Südlichen Hornraben rot ist. Das Verbreitungsgebiet der beiden Arten überschneidet sich in Uganda und Kenia. Hornraben ernähren sich hauptsächlich von großen Insekten und kleinen Wirbeltieren.
Die Nashornvögel kommen in Afrika und Asien vor. Insgesamt gibt es 15 Gattungen mit über 60 Arten, wobei die Artenzahl variieren kann, da einige Unterarten den Status einer Art erhalten haben. Auf beiden Kontinenten kommen etwa gleich viele Arten vor. Thailand und Indonesien haben jeweils 13 Arten, die Philippinen und Malaysia jeweils 10. In Afrika haben Uganda 16 Arten, die Demokratische Republik Kongo 13 und Kamerun 12 Arten. Während die meisten asiatischen Arten Bewohner dichter Wälder sind, gibt es in Afrika neben vielen Regenwaldarten auch viele Arten, die in der Savanne leben.
Von den 15 Gattungen der Nashornvögel kommen 10 Gattungen in Asien und 5 in Afrika vor. Die asiatischen Gattungen sind die Braunhornvögel (Anorrhinus) mit drei Arten, die asiatischen Tokos (Ocyceros) mit drei Arten, die Schwarzhornvögel (Anthracoceros) mit fünf Arten, die Großen Hornvögel (Buceros) mit drei oder vier Arten, der Schildschnabel (Rhinoplax vigil) mit einer Art, die nur auf den Philippinen vorkommenden Tariktikhornvögel (Penelopides) mit 5 Arten, der Nepalhornvogel (Aceros) mit einer Art, die Asiatischen Kehlsack-Hornvögel (Rhabdotorrhinus) mit vier Arten und die Gattung Rhyticeros mit 6 Arten, von denen eine – der Papua-Hornvogel (Rhyticeros plicatus), auf Neuguinea vorkommt. Die Gattung der Langschopf-Hornvögel (Gattung Berenicornis) hat eine Art. Sie ist näher mit der afrikanischen Gattung Horizoceros, die mit vier Arten die afrikanischen Regenwälder bewohnen, verwandt als mit den asiatischen Gattungen.
Neben der Gattung Horizoceros kommen in Afrika die Savannen bewohnenden Tokos (Tockus) mit 10 Arten, die Gattung Lophoceros mit acht Arten, bei denen einige Arten in Regenwäldern, die anderen in der Savanne leben, die Afrikanische Kehlsack-Hornvögel (Bycanistes) mit sechs oder sieben Arten und die Waldhornvögel (Ceratogymna) mit zwei Arten. Die beiden Letzt genannten Gattungen sind überwiegend Bewohner der afrikanischen Regenwälder.
Einige Arten haben sehr große Verbreitungsgebiete. Dazu zählt beispielsweise der Orienthornvogel (Anthracoceros albirostris), der vom Himalaya und den Süden Chinas bis nach Indonesien vorkommt. Auch einige der afrikanischen Tokos, wie beispielsweise der Grautoko (Lophoceros nasutus) oder der Elstertoko (Lophoceros fasciatus), haben sehr große Verbreitungsgebiete. Daneben gibt es eine Reihe von Arten mit sehr kleinen Verbreitungsgebieten. Diese Arten sind vor allem unter den asiatischen Nashornvögeln zu finden, die nur auf einigen Inseln zu finden sind. Sie sind durch Lebensraumveränderungen in besonderem Maße betroffen und entsprechend sind viele Nashornvogelarten mit solch kleinen Verbreitungsgebieten besonders stark bedroht: Der Palawanhornvogel (Anthracoceros marchei) zählt beispielsweise zu den Endemiten Palawans, einer 450 Kilometer langen und bis zu 40 Kilometer breiten philippinischen Insel, die für ihren hohen Grad an Biodiversität und Endemismus bekannt ist. Seine Bestandssituation gilt als gefährdet. Der vom Aussterben bedrohte Suluhornvogel (Anthracoceros montani) kommt dagegen vermutlich nur noch auf der Insel Tawi-Tawi vor, der auf nur noch 502 Brutpaare geschätzte Panayhornvogel (Rhabdotorrhinus waldeni) brütet nur noch auf Panay. Der ebenfalls stark bedrohte Narcondamhornvogel (Rhyticeros narcondami) ist nur auf der Insel Narkondam beheimatet.
Die kleinste Art ist der afrikanische Zwergtoko (Lophoceros camurus) mit 30 cm Körperlänge und einem Gewicht von etwa 110 g. Die größte Art ist der asiatische Doppelhornvogel (Buceros bicornis), mit 1,2 m Körperlänge, wobei der Schnabel etwa 30 cm lang ist, und etwa 3 kg Körpergewicht. Männchen sind tendenziell größer als die Weibchen. Namensgebend für diese Familie ist der große, meist gebogene Schnabel, der außer bei den Tokos einen mehr oder weniger großen wulstigen Aufsatz besitzt. Bei den meisten Arten der Toko-Gattung ist das nashornvogeltypische Horn auf einen wenig auffälligen Schnabelfirst reduziert. Das „Horn“ ist meist hohl oder besteht aus lockerem Knochengewebe, nur beim Schildschnabel (Rhinoplax vigil) ist es massiv (Surapaneni et al. 2025). Beim Doppelhornvogel hat man Hörner gemessen, die etwa 19 Zentimeter lang, über 10 Zentimeter breit und 5 Zentimeter hoch waren. Das Horn dient bei vielen Arten als ein Resonanzkörper für ihre Rufe. Beim Schildschnabel wird das Horn bei Luftkämpfen als Rammbock genutzt. Der schwere Schnabel wird von kräftigen Nackenmuskeln und verschmolzenen Halswirbeln stabilisiert. An Kopf und Hals finden sich nackte, auffallend gefärbte Hautpartien. Bei allen Arten findet sich Drohverhalten, bei dem der Schnabel angehoben und damit der unbefiederte und häufig kontrastreich gefärbte Kehlfleck präsentiert wird. Bei den meisten Arten gibt es nicht nur einen Größenunterschied zwischen Weibchen und Männchen, sondern ein Geschlechtsdimorphismus ist auch in der Färbung des Gefieders, des Schnabels und des Horns vorhanden.
Der lange Schnabel, der lange Hals und die kräftigen Füße erlauben ein breites Nahrungsspektrum, welches aus Früchten, Insekten und kleinen Wirbeltieren besteht. Unter- und Oberschnabel treffen nur an der Spitze genau aufeinander, so dass der Schnabel wie eine Pinzette eingesetzt werden kann. Nashornvögel sind damit in der Lage, mit großer Geschicklichkeit nach Objekten zu greifen. Nashornvögel, die überwiegend von tierischem Protein leben, kommen überwiegend in der afrikanischen Savanne vor. Rotschnabeltoko (Tockus erythrorhynchus) und Monteiro-Toko (Tockus monteiri) ernähren sich dabei fast ausschließlich von tierischer Kost. Bei allen anderen Nashornvogelarten überwiegt eine Ernährung mit Früchten, Beeren und Samen. Doch auch hier wird tierische Nahrung nicht vermieden und bei einigen Arten wie dem Orienthornvogel (Anthracoceros albirostris) und dem Silberwangenhornvogel (Bycanistes brevis) ist beobachtet worden, dass sie Vögel bis Taubengröße erbeuten. Viele insektenfressende Nashornvogelarten folgen auch anderen Tierarten, die während ihrer Nahrungssuche Insekten aufschrecken. Vom Von-der-Decken-Toko (Tockus deckeni) und Östlichem Gelbschnabeltoko (Tockus flavirostris) ist sogar bekannt, dass sie mit Zwergmangusten kooperieren. Die beiden Toko-Arten fangen die Insekten (insbesondere Heuschrecken), die diese in Gruppen lebenden Mangusten bei ihrer Nahrungssuche aufscheuchen. Die Zwergmangusten profitieren, weil beide Tokos vor Beutegreifern aus der Luft warnen.
Nashornvögel brüten meistens in Baumhöhlen. Allen Arten ist gemeinsam, dass sich das Weibchen in der Bruthöhle bis auf einen schmalen Spalt einmauert. Meist werden dafür Exkremente und Nahrungsreste, selten auch Lehm genutzt. Dadurch ist der Nachwuchs zwar weitgehend vor Nesträubern geschützt, allerdings ist das Männchen über eine lange Zeit allein für die Nahrungsversorgung zuständig. Beim Silberwangenhornvogel (Bycanistes brevis), bei dem das Weibchen bis zu 138 Tage in der Bruthöhle bleibt, bringt das Männchen täglich etwa 360 Gramm Früchte zum Nest, die das Männchen im Schlund oder Schnabel herbeiträgt. Das Weibchen hält die Bruthöhle sauber, indem es durch den schmalen Spalt kotet. Auch die Jungvögel zeigen ein solches Verhalten, sobald sie eine hinreichende Größe erreicht haben. Bei einer Reihe von Arten verlässt das Weibchen vor dem Flüggewerden der Jungvögel die Bruthöhle und versorgt den Nachwuchs gemeinsam mit dem Männchen mit Nahrung. Bei anderen Arten verbleibt das Weibchen bis zum Flüggewerden des Nachwuchses in der Bruthöhle. Der Nachwuchs ist zu dem Zeitpunkt, zu dem er die Nisthöhle verlässt, flugfähig. Er kehrt nicht in die Bruthöhle zurück. Das Weibchen durchläuft gewöhnlich während der Brut die Mauser. Bei einigen Arten wie beispielsweise den Tokos vermausert das Weibchen das Großgefieder gleichzeitig und ist deshalb zeitweise flugunfähig. Bei anderen Arten wird das Großgefieder nacheinander vermausert, die Weibchen behalten dabei ihre Flugfähigkeit.
Die frühesten fossilen Nachweise von Nashornvögeln stammen erst aus dem mittleren Miozän von Marokko und Uganda. Das in Marokko gefundene Fossil gehört zu einer ausgestorbenen Art der Hornraben (Bucorvus brailloni), während die in Uganda lebende Art zu den Tokos gehört. Dass Nashornvögel im Miozän auch außerhalb ihres modernen Verbreitungsgebietes heimisch waren, belegt ein Fossil aus Bulgarien: Euroceros bulgaricus. Ansonsten sind die fossilen Nashornvögel schlecht dokumentiert. Da sie eine enge Verwandtschaft mit den Baumhopfen und Wiedehopfen teilen und diese schon im Eozän nachgewiesen sind, dürfte die Evolutionsgeschichte der Nashornvögel wesentlich älter sein, als fossil dokumentiert (Brunet 1971, Boev & Kovachev 2007, Riamon et al. 2021).
Innerhalb der Coraciimorphae finden sich noch zwei weitere Ordnungen, die wir in der nächsten Episode kennenlernen werden.
Literatur
Boev, Z., Kovachev, D. (2007). Euroceros bulgaricus gen. nov., sp. nov. from Hadzhidimovo (SW Bulgaria) (Late Miocene) – the first European record of Hornbills (Aves: Coraciiformes). – Geobios, 40: 39–49.
Brunet, J. (1971). Oiseaux miocènes de Beni Mellal (Maroc); un complément à leur étude. Notes Mem. Serv. geol. Maroc, 31 (237): 109–111.
Del Hoyo, J. (2020): All the Birds of the World. Lynx Nature Books
Del Hoyo, J.; Elliot, A.; Sargatal, J., eds. (2001): Handbook of the Birds of the World, volume 6, Mousebirds to Hornbills, Lynx Edicions, Barcelona.
Houde, P., Olson, S. L. (1992). A radiation of coly-like birds from the early Eocene of North America (Aves: Sandcoleiformes new order). Natural History of Museum of Los Angeles County, Science Series 36: 137-160.
Johansson, U.S., Ericson, P.G.P. (2005). A re-evaluation of basal phylogenetic relationships within trogons (Aves: Trogonidae) based on nuclear DNA sequences. Journal of Zoological Systematics and Evolutionary Research 43,166–173.
Kristoffersen, A.V. (2002). An early Paleogene trogon (Aves: Trogoniformes) from the Fur Formation, Denmark. Journal of Vertebrate Paleontology, 22(3): 661–666. doi:10.1671/0272-4634(2001)022[0661:AEPTAT]2.0.CO;2
Ksepka, D.T., Clarke, J. A. (2009). Affinities of Palaeospiza bella and the phylogeny and biogeography of mousebirds (Coliiformes). The Auk 126: 245-259. doi: 10.1525/auk.2009.07178
Ksepka, D.T., Clarke, J. A. (2010). New fossil mousebird (Aves: Coliiformes) with feather preservation provides insight into the ecological diversity of an Eocene North American avifauna. Zoological Journal of the Linnean Society 160: 685-706. doi: 10.1111/j.1096-3642.2009.00626.x
Ksepka, D.T., Stidham, T.A., Williamson, T.E. (2017). Early Paleocene landbird supports rapid phylogenetic and morphological diversification of crown birds after the K–Pg mass extinction. PNAS 114: 8047-8052. doi: 10.1073/pnas.1700188114
Martín-Platero, A.M. et al. (2006). Characterization of antimicrobial substances produced by Enterococcus faecalis MRR 10-3, isolated from the uropygial gland of the hoopoe (Upupa epops)., Appl Environ Microbiol. 72 (6): S. 4245–4249
Martín-Vivaldi, M. et al. (2010). Antimicrobial chemicals in hoopoe preen secretions are produced by symbiotic bacteria., Proc Biol Sci. 277(1678):123-30
Mayr, G. (1999). A new trogon from the Middle Oligocene of Céreste, France. The Auk 116(2): 427–434. doi: 10.2307/4089376
Mayr, G. (2000). A new mousebird (Coliiformes: Coliidae) from the Oligocene of Germany. Journal für Ornithologie 141: 85-92. doi: 10.1007/BF01651775
Mayr, G. (2001). New specimens of the Middle Eocene fossil mousebird Selmes absurdipes Peters 1999. Ibis 143: 427-434. doi: 10.1111/j.1474-919X.2001.tb04943.x
Mayr, G. (2005). A new Eocene Chascacocolius-like mousebird (Aves: Coliiformes) with a remarkable gaping adaptation. Organisms Diversity and Evolution 5: 167-171. doi: 10.1016/j.ode.2004.10.013
Mayr, G. (2009). Paleogene Fossil Birds. Heidelberg: Springer
Mayr, G. (2009a). A well-preserved second trogon skeleton (Aves, Trogonidae) from the middle Eocene of Messel, Germany. Palaeobiodiversity and Palaeoenvironments. 89 (1–2): 1–6. doi: 10.1007/s12549-009-0001-9
Mayr, G. (2010). Mousebirds (Coliiformes), parrots (Psittaciformes), and other small birds from the late Oligocene/early Miocene of the Mainz Basin, Germany. Neues Jahrbuch für Geologie und Paläontologie – Abhandlungen 258: 129-144. doi: 10.1127/0077-7749/2010/0089
Mayr, G. (2013). Late Oligocene mousebird converges on parrots in skull morphology. Ibis 155: 384-396. doi: 10.1111/ibi.12034
Mayr, G. (2015). A new specimen of the Early Eocene Masillacolius brevidactylus and its implications for the evolution of feeding specializations in mousebirds (Coliiformes). Comptes Rendus Palevol 14: 363-370. doi: 10.1016/j.crpv.2015.05.007
Mayr, G. (2016): Avian Evolution: The Fossil Record of Birds and its Paleobiological Significance. TOPA Topics in Paleobiology.
Mayr, G. (2018). New data on the anatomy and palaeobiology of sandcoleid mousebirds (Aves, Coliiformes) from the early Eocene of Messel. Palaeobiodiversity and Palaeoenvironments 98: 639-651. doi: 10.1007/s12549-018-0328-1
Mayr, G., Mourer-Chauviré, C. (2004). Unusual tarsometatarsus of a mousebird from the Paleogene of France and the relationships of Selmes Peters, 1999. Journal of Vertebrate Paleontology 24: 366-372. doi: 10.1671/1970
Mayr, G., Peters, D. S. (1998). The mousebirds (Aves: Coliiformes) from the Middle Eocene of Grube Messel (Hessen, Germany). Senckenbergiana lethaea 78: 179-197. doi: 10.1007/BF03042768
Mayr, G., et al. (2019). Skeletons from the early Oligocene of Poland fill a significant temporal gap in the fossil record of upupiform birds (hoopoes and allies). Historical Biology, 32(9), 1163–1175. https://doi.org/10.1080/08912963.2019.1570507
Mayr, G. et al. (2023). Narrow-beaked trogons from the early Eocene London Clay of Walton-on-the-Naze (Essex, UK). J Ornithol, doi: 10.1007/s10336-023-02071-x
Mlíkovský, J., Göhlich, U. (2000). A new wood-hoopoe (Aves: Phoeniculidae) from the early Miocene of Germany and France. Acta Societatis Zoologicae Bohemicae. 64. 419-424.
Olson, S.L. (1975). Paleornithology of St. Helena Island, south Atlantic Ocean, Smithsonian Contributions to Paleobiology 23
Peters, D.S. (1999). Selmes absurdipes, new genus, new species, a sandcoleiform bird from the oil shale of Messel (Germany, Middle Eocene). Smithsonian Contributions to Paleobiology 89: 217-222.
Riamon, S.; Pickford, M.; Senut, B.; Louchart, A. (2021). Bucerotidae from the early Miocene of Napak, Uganda (East Africa): The earliest hornbill with a modern-type beak. Ibis. 163 (2): 715–721.
Sangster, G., Gregory, S.M.S., Dickinson, E.C. (2023). Apalodermatinae, a new family-group name for the African trogons (Trogonidae), with clarification of the authorship of Harpactini. Avian Systematics 2023 2 (3): 45–52
Saupe, E.E., et al. (2019). Climatic shifts drove major contractions in avian latitudinal distributions throughout the Cenozoic. PNAS 116: 12895-12900. doi: 10.1073/pnas.1903866116
Surapaneni, V. et al. (2025). The helmeted hornbill casque is reinforced by a bundle of exceptionally thick, rod‐like trabeculae. Annals of the New York Academy of Sciences. 1544. 78-91. 10.1111/nyas.15254.
Winkler, D. W., Billerman, S. M., Lovette, I. J. (2015): Bird Families of the World: A Guide to the Spectacular Diversity of Birds. Lynx Edicions, Barcelona.
Winkler, D. W., Billerman, S. M., Lovette, I. J. (2020a). Mousebirds (Coliidae), version 1.0. In Birds of the World (S. M. Billerman, B. K. Keeney, P. G. Rodewald, and T. S. Schulenberg, Editors). Cornell Lab of Ornithology, Ithaca, NY, USA. https://doi.org/10.2173/bow.coliid1.01
Winkler, D. W., Billerman, S. M., Lovette, I. J. (2020b). Cuckoo-roller (Leptosomidae), version 1.0. In Birds of the World (S. M. Billerman, B. K. Keeney, P. G. Rodewald, and T. S. Schulenberg, Editors). Cornell Lab of Ornithology, Ithaca, NY, USA. https://doi.org/10.2173/bow.leptos1.01
Winkler, D. W., Billerman, S. M., Lovette, I. J. (2020c). Trogons (Trogonidae), version 1.0. In Birds of the World (S. M. Billerman, B. K. Keeney, P. G. Rodewald, and T. S. Schulenberg, Editors). Cornell Lab of Ornithology, Ithaca, NY, USA. https://doi.org/10.2173/bow.trogon1.01
Winkler, D. W., Billerman, S. M., Lovette, I. J. (2020d). Hoopoes (Upupidae), version 1.0. In Birds of the World (S. M. Billerman, B. K. Keeney, P. G. Rodewald, and T. S. Schulenberg, Editors). Cornell Lab of Ornithology, Ithaca, NY, USA. https://doi.org/10.2173/bow.upupid1.01
Winkler, D. W., Billerman, S. M., Lovette, I. J. (2020e). Woodhoopoes and Scimitarbills (Phoeniculidae), version 1.0. In Birds of the World (S. M. Billerman, B. K. Keeney, P. G. Rodewald, and T. S. Schulenberg, Editors). Cornell Lab of Ornithology, Ithaca, NY, USA. https://doi.org/10.2173/bow.phoeni2.01
Winkler, D. W., Billerman, S. M., Lovette, I. J. (2025f). Hornbills (Bucerotidae), version 1.1. In Birds of the World (S. M. Billerman, B. K. Keeney, P. G. Rodewald, and T. S. Schulenberg, Editors). Cornell Lab of Ornithology, Ithaca, NY, USA. https://doi.org/10.2173/bow.bucero1.01.1
Zelenkov, N.V., Dyke, G.J. (2008). The fossil record and evolution of mousebirds (Aves: Coliiformes). Palaeontology 51: 1403-1418. doi: 10.1111/j.1475-4983.2008.00814.x
