
zu Teil 7:
Einführendes zum Gehirn
Als Gehirn wird bei Wirbeltieren und einigen Wirbellosen der im Kopf gelegene Teil des Zentralen Nervensystems (ZNS) bezeichnet (als weiterer Bestandteil des zentralen Nervensystems der Wirbeltiere gilt das Rückenmark). Unser Gehirn verarbeitet Sinneswahrnehmungen und koordiniert das Verhalten. Die Länge aller Nervenbahnen zusammengenommen beträgt ungefähr 5,8 Millionen Kilometer (145-fache des Erdumfangs). Das Volumen des Gehirns beträgt bei erwachsenen Männern durchschnittlich 1,27 Liter, bei Frauen 1,13 Liter; das Hirngewicht erwachsener Männer beträgt ungefähr 1400 g, bei gleichgroßen Frauen ist es 100 g leichter (ALLEN, DAMASIO & GRABOWSKI 2002, RUSHTON 1992).
Einteilung des Gehirns bei Menschen (Quelle)
Vereinfachend lässt sich das Gehirn nach in Großhirn (Telencephalon) mit Hirnrinde (Cortex), Kleinhirn (Cerebellum, Metencephalon), Zwischenhirn (Diencephalon), welches unter anderem die Hypophyse und den Thalamus beinhaltet und Hirnstamm (Truncus cerebri) mit Mittelhirn (Mesencephalon) und Nachhirn (Myelencephalon) eingeteilt.
Das Großhirn –besteht aus 2 Hemisphären, welche durch das Corpus Callosum (Balken) miteinander verbunden sind. Die Oberfläche des Großhirns ist die Großhirnrinde, oder auch cerebraler Cortex genannt. Die Großhirnrinde ist die größte Struktur des menschlichen Gehirns (ausgebreitet etwa so groß wie die Titelseite einer Tageszeitung) und wird in Rindenfelder unterteilt. Die Großhirnrinde besteht aus 16 Milliarden Nervenzellen und etwa 2-4 mm dick. Unter der Rinde verlaufen die Nervenfasern. Die Neuronen sind bei lebenden Gehirnen rosa, färben sich aber nach dem Tod grau, man spricht daher von grauer Substanz. Die Nervenfasern bezeichnet man entsprechend ihrer Farbe als weiße Substanz. Die Rindenfelder sind eingeteilt in den Frontallappen (Stirnseite des Kopfes), den Parietallappen (am Scheitel des Kopfes), den Temporallappen (an des Seiten des Kopfes) und den Occipitallappen (im hinteren Bereich des Kopfes, vgl. Abb. 1).
Der Frontallappen ist das oberste Kontrollzentrum für situationsbezogenes Handeln sowie die Verarbeitung emotionaler Prozesse. Er empfängt die verarbeiteten sensorischen Signale, verknüpft sie mit Gedächtnisinhalten und emotionalen Bewertungen und initiiert auf dieser Basis Handlungen.
Der Parietallappen ist an der Körperwahrnehmung (Wahrnehmung von Druck, Temperatur, Vibration, Oberflächenbeschaffenheit) sowie dem räumlichen Denken beteiligt.
Der Temporallappen ist mit dem auditorischen Kortex für das Hörvermögen verantwortlich und beinhaltet zudem den Hippocampus. Dieser ist mit dafür verantwortlich, wie Fakten und autobiografische Ereignisse mittel- bis langfristig im Gedächtnis gespeichert werden. An der Spitze des Hippocampus befindet sich die Amygdala, welche Informationen emotional bewertet und damit ebenfalls mit dafür verantwortlich ist, welche Informationen langfristig gespeichert werden.
Der Occipitallappen beinhaltet das primäre und sekundäre Sehzentrum und erlaubt es somit, die Welt zu betrachten und zu erkennen.
Das Kleinhirn – oder Cerebellum – befindet sich leicht unterhalb des Großhirns und ist insbesondere an den Prozessen für Gleichgewicht, Bewegung und Koordination beteiligt. Damit ist es besonders bei Lernprozessen mit motorischen Reaktionen wichtig.
Das Zwischenhirn – Diencephalon – besteht aus dem Thalamus, Hypothalamus, Subthalamus und Epithalamus. Der Thalamus erhält sensorische Informationen vom peripheren Nervensystem und leitet diese an das Großhirn weiter. Er ist damit eine Schnittestelle, in der die sensorischen Informationen des Gehirns zusammenlaufen. Der Hypothalamus steuert zahlreiche körperliche und psychische Vorgänge und bildet zusammen mit der Hypophyse das Bindeglied zwischen Hormon- und Nervensystem.
Der Hirnstamm – Truncus cerebri – befindet sich an der Basis des Gehirns und besteht aus Mittelhirn, Brücke und Nachhirn, welches auch den Übergang zum Rückenmark bildet. Der Hirnstamm steuert die automatisch bzw. reflexartig ablaufenden Vorgänge (Herzschlag, Atmung, Regulierung der Körpertemperatur, Schluck- und Hustenreflex) des menschlichen Körpers.
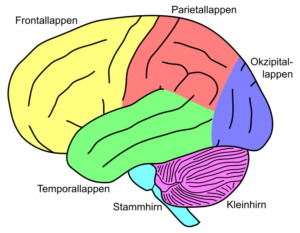
Abb. 1: die Großhirnrinde des Menschen mit Kleinhirn und Stammhirn
Gedankenlesen?
Dass irgendwelche Esoterik-Damen und Orakel durch das Schauen in eine Glaskugel oder unsere Handflächen meinen die Zukunft vorhersagen zu können, das bestreitet jeder logisch und vernünftig denkende Mensch (hoffe ich). Aber wie sieht es denn mit unseren Neurowissenschaftlern aus? Können die wenigstens unsere Gedanken lesen? Das mag vielleicht banal klingen, aber mehrere Massenmeiden stellen die Behauptung auf. Das PM-Magazin titelt 2013: „Ich weiß was du denkst – die Kunst des Gedankenlesens hat sich vom Zirkusspektakel zum Topthema der Hirnforschung entwickelt.“ Das Handelsblatt 2011 hatte schon Zukunftsvisionen: „Gehirnscan – Fortschritte beim Gedankenlesen“ (zitiert in BECK 2014: S. 15). Um Gehirne bei der Arbeit zu beobachten (Aufgabe des Gehirns ist übrigens das Denken und das tun Gehirne immer, auch wenn wir manches als Schwachsinn bezeichnen – unsere Nervenzellen sind immer aktiv) nutzt man komplexe Messgeräte – und verfahren, so die Magnetresonanztomographie (MRT) und Elektroenzephalographie (EEG). Bei einem EEG werden einem mehrere Elektroden auf den Kopf platziert und die elektrischen Felder der Nervenzellen im Gehirn gemessen (Abb. 2). Da sich in unserem Gehirn 80 Milliarden (80.000.000.000) Nervenzellen befinden, ist das erzeugte Feld stark genug, um es von außen durch die Schädeldecke hindurch messen kann (ich hatte mal für eine Untersuchung in der Uni so ein EEG gemacht, es ist harmlos, macht aber die Frisur kaputt). Der Vorteil dieser Messungen der Hirnströme ist, dass die Forscher „live“ dabei sind, wenn die Nervenzellen ihre Impulse aussenden. Das EEG findet z. B. Anwendung bei Epilepsie, Schlaffunktionen, Koma oder auch bei der Messung allgemeiner Hirnaktivität. Jedoch ist eine genauere Lokalisation der Erzeugung dieser Ströme nicht bzw. nur sehr ungenau bestimmbar. Will man untersuchen, welche Gehirnregionen bei bestimmten Reizen oder Gedanken besonders beansprucht werden, so nutzt man die funktionelle Magnetresonanztomographie (fMRT), umgangssprachlich spricht man von „Hirnscans“. Ohne auf die komplizierten Rechenvorgänge einer fMRT eingehen zu wollen, müssen wir an dieser Stelle nur eines Wissen: Es handelt sich um ein bildgebendes Verfahren, welche es möglich macht Durchblutungsänderungen von Hirnarealen sichtbar zu machen. Werden gewisse Hirnareale stärker beansprucht, müssen sie mehr durchblutet werden, um eine Sauerstoffversorgung der Zellen zu gewährleisten. Stärker durchblutete Areale werden in einer bildlichen Darstellung farblich hervorgehoben. Daraus ist übrigens nicht zu schließen, dass die nicht farblich markierten Bereiche nicht aktiv sind, sie sind nur weniger aktiv als die farblich hervorgehobene Hirnregion. Außerdem können die besten Hirnscans nicht die wahren Gedankenvorgänge im Gehirn aufzeichnen, man kann nur feststellen, wo im Gehirn aktiv etwas passiert, was man aber genau denkt (ob es nun eine rote Blume oder eine blaue Blume ist), kann man nicht feststellen. Das hat auch nicht zuletzt damit zu tun, dass unsere Gedanken (bzw. die Nervenimpulse) viel schneller ablaufen, als eine fMRT diese aufzeichnen kann, man misst halt nur Durchblutungsmuster. Damit ist auch das Messverfahren als solches indirekt: man misst nicht den Gedanken als solches, sondern eine stärkere Durchblutung.
Der Science-Journalist Greg Miller vergleicht das ausspionieren von Neuronen mit dem Kalten Krieg: „Das Benutzen von fMRT zum Ausspionieren von Neuronen ist vergleichbar mit der Benutzung von Satelliten aus dem Kalten Krieg, um damit Menschen auszuspionieren: man kann nur großflächige Aktivität sichtbar machen.“ (MILLER 2008)
Damit ist nichts weiter gemeint, als die Tatsache, dass die Neuronen so schnell arbeiten (innerhalb weniger Sekunden können Millionen von Neuronen jeweils hunderttausende von Nervenimpulsen generieren), dass wir nicht in der Lage sind, den gesamten Prozess mit einem fMRT sichtbar zu machen, sondern nur einen Bruchteil. Dies schränkt aber auch die Interpretationsmöglichkeiten stark ein.
Wer also behauptet, Neurowissenschaftler können ihre Gedanken lesen – und leider gibt es nicht wenige Markenfuzzies, die das behaupten; es gibt sogar ein Neuromarketing – so ist diese Aussage deutlich übertrieben. Hirnforscher messen die Aktivität des Gehirns, vom Wissen über den Gedankeninhalt sind sie meilenweit entfernt.
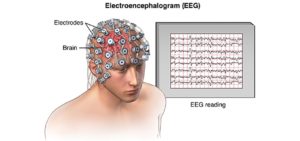
Abb. 2: links EEG, recht fMRT-Aufnahme
Rattenhirn = Menschenhirn?
Das meiste, was wir über Gehirne und Hormone wissen, wissen aus Tierversuchen. Sie liefern uns wertvolle Grunderkenntnisse, wie die Neurophysiologie funktioniert. Doch sollte man vorsichtig damit sein Ergebnisse direkt von z. B. einem Rattenhirn auf ein Menschenhirn zu übertragen. Denn neben vielen natürlichen Gemeinsamkeiten gibt es auch bedeutende Unterschiede. HINES (2004, S. 82) hebt hervor, dass ein Penis ein Penis ist und sowohl bei männlichen Ratten wie auch bei Männern dieselbe Funktion hat. Ein Nagerhirn lässt sich jedoch schon alleine wegen der Relation der Gehirngröße zur Körpermasse, sowie der Hirnrinde nicht mit dem menschlichen Gehirn vergleichen. Auch haben die Hirnregionen unterschiedliche und beim Menschen auch komplexere Aufgaben und Strukturen (JORDAN-YOUNG 2010, S. 48)
WALLEN (2005) geht davon sogar aus, dass die Daten von Hormonstudien, die man aus Ratten gewonnen hat, wenig darüber Auskunft geben, was tatsächlich beim Menschen vor sich geht. So gibt es Unterschiede wann, wo und wie die Hormone bei Menschen und Ratten wirken. Das gilt übrigens nicht nur für Ratten, sondern natürlich auch für andere Versuchstiere. Anders als bei Tieren sind Hormone bei Menschen weniger relevant für die Determination des Verhaltens aufgrund unserer komplexen Gehirne, wodurch man bei Übertragungen der Ergebnisse von Tierversuchen auf menschliches Verhalten sehr vorsichtig sein sollte (HALPERN 2012, S. 183).
RIPPON (2010) kommentiert in ihrem Vortrag „Sexing the Brain“, dass wenn man sich für das geschlechtsspezifische Verhalten von Fruchtfliegen, Zebrafischen oder Mäusen interessiert, sind die zu bewerkstelligenden Experimente eindeutig. Bei Fragen, die sich auf das Verhalten von Menschen konzentrieren, wird die Sache jedoch komplizierter. Um dies zu vereinfachen werden dass die Ergebnisse, die man bei Zebrafischen gefunden hat direkt auf den Menschen übertragen. Das sind laut RIPPON (2010) typische „brain blooper“ (Hirnpatzer), da solche Ergebnisse nicht direkt übertragbar sind.
Die plastische, also anpassungsfähige, Funktionsweise des Gehirns hat auch der sowjetische Physiologe Iwan Pawlow erkannt. Die meisten dürfte dieser Name vertraut sein, wenn es um das Thema Konditionierung geht. Doch seine Studien und Erkenntnisse reichen wesentlich weiter. Pawlow hatte sich mit den Funktionen des Gehirns auseinandergesetzt. Er entwickelte die Theorie der drei Systeme der höheren Nerventätigkeit: die unbedingten und bedingten Reflexe, die bei Menschen und allen anderen Tierarten vorkommen und das Signalsystem der Sprache, welches nur dem Menschen eigen ist.
„Der Begriff Reflex bezeichnet eine wissenschaftliche Vorstellung, der zufolge ein bestimmter Reiz von außen oder aus dem inneren Organsystem des Tieres eine bestimmte Wirkung an einem Nervenrezeptor (Augen, Nasen, Ohren, Haut, Schleimdrüse etc.) hervorruft. Diese Wirkung wird dann in einen nervalen Prozess umgewandelt, d. h. in eine Nervenerregung. Die Erregung wird entlang bestimmter Nervenfasern zum zentralen Nervensystem weitergeleitet, wo schon vorher bestehende Verbindungen genutzt oder neue hergestellt werden.“ (WELLS 1989, S. 72)
„Die Großhirnrinde beinhaltet demgegenüber den Apparat für die ständige Anpassung an täglich wechselnde Umweltbedingungen. Das kortikale Signalsystem ist selbst ständig in einem Fließzustand, der dem der weniger beständigen Umweltbedingungen entspricht. Das tägliche Leben erfordert sehr fein verzweigte und spezialisierte Wechselbeziehungen zwischen dem Tier und seiner Umwelt. Die Signalfunktion der Hemisphären macht es möglich, dass eine Vielzahl natürlicher Reize vorübergehend die relativ kleine Anzahl grundlegender Stimuli ersetzt, die entscheidend für die angeborenen Reflexe sind. Einzig durch diese Signaltätigkeit der Großhirnhemisphären wird eine genau abgestimmte und empfindliche, aber dennoch sich ständig ändernde Anpassung des Organismus an die Umwelt erzielt.“ (WELLS 1989, S. 74)
„‚Reflexe und Instinkte sind also gleichermaßen natürliche Reaktionen des Organismus auf bestimmte Reize; daher ist es auch nicht nötig, sie durch unterschiedliche Bezeichnungen voneinander zu scheiden. Der Begriff Reflex ist vorzuziehen, da er von Anfang an in streng wissenschaftlichen Sinne verwendet worden ist.‘ Die sogenannten Instinkte sind nach Pawlow angeborene Reflexketten.“ (zitiert in Wells 1989, S. 92)
Die verschiedenen Signalsysteme arbeiten streng zusammen:
„Von Geburt an werden die angeborenen Reflexe von Tier und Mensch gleichermaßen untrennbar mit erworbenen und zeitweiligen Signalen verflochten. Nach Pawlow stehen als das erste und zweite System nicht nur in enger Beziehung zueinander, sondern sie sind voneinander abhängig. Jede Äußerung eines ‚Instinkts‘ – oder jede Reflexkette – bewirkt gleichzeitig eine noch engere Verflechtung mit den sensorischen Signalen. Das Ergebnis des engen Zusammenwirkens des angeborenen und erworbenen Systems der höheren Nerventätigkeit ist eine Verschmelzung von ‚Instinkten‘ und sensorischen Signalen zu verschmolzenen Reflexen. Durch Wiederholung werden diese verschmolzenen Reflexe zu Systemen verknüpft und damit zu dem, was Pawlow als dynamische Stereotype bezeichnete – oder was auf Ebene der Beschreibung als Verhaltensmuster oder Gewohnheit bekannt ist.“ (WELLS 1989, S. 106)
Das klingt im ersten Moment etwas komplex, ist aber im Grunde genommen recht einfach und wird in jedem Biologie-Schulbuch zitiert: Pawlows Konditionierung vom Hund. Wird einem Hund ein unbedingter Reiz (Futter) präsentiert, zeigt dieser einen angeborenen (unbedingten) Reiz (Speichelfluss). Auf das Läuten eines indifferenten, neutralen reiz (Läuten einer Glocke) kam es zu keiner Reaktion. Kombiniert man beide Reize (Unbedingt: Futter, indifferent: Glocke), reagiert der Hund mit Speichelfluss. Nach mehrmaligem Wiederholen dieser Reizpräsentation, reagiert der Hund schon auf das Glockenläuten mit Speichelfluss. Diese Reaktion nennt Pawlow bedingten Reflex. Entscheidend ist hier, dass ein vorher neutraler Reiz eine Reaktion hervorruft, die vorher nur durch einen unbedingten Reiz ausgelöst wurde. Diese Kombination von unbedingten und bedingten Reizen wie Reflexen ist nach Pawlow grundlegend für das Verhalten von Tieren, aber auch Menschen. Auch wir haben diese angeborenen, unbedingten Reflexe, z. B. Speichelreflex, Knie-Sehnen-Reflex, Greifreflex usw. usf. Und auch diese unbedingten Reflexe werden bei uns verknüpft mit dem Lernen aus Erfahrung. Aber das menschliche Verhalten erschöpft sich nicht nur auf diese zwei Signalsysteme. Pawlow und seine Schule (Bykow, Lruija, Wygotski, Anochin, Leontijew etc.) erkannten, dass es bei Menschen noch ein weiteres Signalsystem gibt: das Signalsystem der Sprache.
„Als Signale gleichen Wörter einerseits allen anderen bedingten Reizen. Sie unterliegen den allgemeinen Gesetzen der Ausbildung und der Löschung bedingter Reflexe und der Erhöhung ihrer Spezifität (…) Andererseits unterscheiden sich die Wörter jedoch qualitativ von allen anderen bedingten Reizen. Der Unterschied besteht in ihrer Abstraktheit. (…) Im Vergleich zu der ungeheureren Flexibilität der Reaktionen aber, die dem Menschen durch das Sprachsystem zur Verfügung stehen, sind die bedingten Reflexe auf sensorische Reize bei Tieren und Säuglingen relativ grob und primitiv. Das wird vollends deutlich, wenn wir uns klar machen, dass das menschliche Denken vor allem in Selbstkommunikation mittels der Sprache besteht und dass die Arbeit und Produktion, Landwirtschaft und Technik, ebenso wie Wissenschaft und Kunst von der Existenz der Sprache oder des Signalwegs der Wörter abhängen. In Begriffen der historischen Entwicklung heißt das, dass sich das System der bedingten Reflexe der Sprache in und durch und mit dem gesellschaftlichen Leben und der Arbeit, der Produktion, der Landwirtschaft, der Kunst und Wissenschaft entwickelt hat, so dass das dritte System gleichzeitig sowohl das Produkt dieser menschlichen Tätigkeit als auch eine Bedingung ihrer Entwicklung ist.“ (WELLS 1989, S. 108 – 109)
„Auf dieser Weise kann der Mensch denken, bevor er handelt, d. h. er kann mittels logisch bestimmter Wortassoziationen stellvertretend verschiedene Handlungsabläufe verfolgen und erst handeln, wenn er den geeignetsten ausgewählt hat. Gezieltes Handeln dieser Art ist nur dem Menschen möglich, da es den Denkprozess voraussetzt, der wiederrum abhängt von der höheren Nerventätigkeit des Sprachsystems und der sensorischen Systeme.“ (WELLS 1989, S. 113)
Die Erkenntnisse von Pawlow und seiner Schüle liefern Hinweise darauf, warum unsere Gehirne so plastisch sind (siehe Kapitel Gehirn und Umwelt) und weshalb es schwer ist, von tierischen Hirnvorgängen auf menschliche zu schließen. Als Menschen sind wir soziale Wesen in der Lage uns gegenseitig und unser Umfeld zu beeinflussen, unsere Tätigkeit wird geformt über unsere Sprache, die qualitativ höherwertiger ist als jede Kommunikation im Tierreich, inklusive der Menschenaffen. Wir wissen z. B., dass Menschenaffen hoch intelligent sind: sie können Werkzeuge gebrauchen, bauen sich Schlafnester, sie sind emotional und man kann ihnen sogar unter menschlicher Obhut die Zeichensprache wie auch den Umgang mit Feuer beibringen. So erstaunlich diese Leistungen auch sein mögen, müssen jedoch zwei Aspekte mit einbezogen werden:
- Hat der Affe kein Plan vom „Werkzeug“ im Kopf. Er ist intelligent genug einen Stein zu sehen und damit eine Nuss zu knacken. Aber das tut er nur, wenn er das Bedürfnis hat, eine Nuss zu essen. Nie ist ein Affe auf die Idee gekommen den Stein (oder ein anderes Werkzeug) „effizienter“ zu bearbeiten, wie einen Faustkeil oder diesen wenigstens mitzunehmen. Er nimmt irgendeinen Stein, wenn er das Bedürfnis hat eine Nuss zu essen. Aber kein Affe steht morgens auf und plant im Voraus Nüsse und Steine zu sammeln, wenn er gerade mit der fellpflege beschäftigt ist, weil dies der Affe nur tut, wenn er das unmittelbare Bedürfnis verspürt, wenn er Nüsse sieht. Findet der Affe nun aber in der Nähe keinen Stein oder einen Stock, um die Nuss zu knacken, wird er sich auch nicht auf die Suxche nach einem machen, erst nicht einen produzieren. Weiterhin fehlt dem Affen die Abstraktion des Begriffes Werkzeug (oder Nuss), der bei Menschen mittels der Sprache vermittelt wird (Menschenaffen sind übrigens weder genetisch, noch anatomisch in der Lage zu sprechen). Und genau das bestimmt den qualitativen Unterschied zwischen dem „Arbeitstier“ Mensch und dem nicht arbeitenden Affen, weil erster nicht nur seine unmittelbaren Bedürfnisse befriedigt, die ihn gerade jetzt Triggern, sondern schon in die Zukunft plant (auch ein Plan des Werkzeuges in seinem Kopf hat), obwohl er nicht mal sicher sein kann den nächsten Tag zu erleben.
- Menschenaffen kann man sicherlich – in gewissen Grenzen – einige komplexe Dinge beibringen, wie Zeichensprache. Doch der entscheidende Punkt ist, dass dies der Affe nur unter Obhut des Menschen erlernt. Bisher ist nicht bekannt, dass ein Schimpanse, Bonobo oder Gorilla irgendwo im afrikanischen Urwald auf die Idee kam, Werkzeuge herzustellen (statt nur zu benutzen), schon gar die Zeichensprache zu lernen oder mit dem Feuer umzugehen.
Diese Aspekte machen die Einzigartigkeit des Menschen aus (und das ist damit gemeint, wenn Friedrich Engels die Bedeutung der Arbeit für die Menschwerdung des Affen hervorhebt).
Gehirn und Umwelt
Ein bahnbrechender Durchbruch zum Verständnis über unser Gehirn gehört die Erkenntnis, dass unser Gehirn plastisch und anpassungsfähig ist, also auf Umweltveränderungen reagiert. Was heißt das konkret? BECK (2014) fasst zusammen:
„Die Funktion des Gehirns bedingt das Verhalten. So entstehen soziale Strukturen, die wieder auf das Gehirn zurückwirken. Das Gehirn ist so plastisch, dass es sich im Laufe der Zeit in diesen Strukturen besser zurechtfindet. Seine Architektur passt sich der Umwelt an. Durch diese ständige Rückkopplung zwischen Gehirn und Umwelt ist der Aufbau des Gehirns gleichzeitig Abbild der Umgebung und Grund für unser Verhalten.“ (BECK 2014, S. 91-92)
Wie anpassungsfähig Gehirne sind zeigt sich auch in einer Studie von JOEL (2011, 2012) dass Umweltfaktoren wie pränataler und postnataler Stress, Drogen, Erziehung oder der Verlust der Mutter mit dem Gehirn in komplexer Weise interagieren. Und um die Dinge zu verkomplizieren interagieren die unterschiedlichen Bereiche des Gehirns mit Hormonen, Genen und Umweltfaktoren auf unterschiedliche Weise (MCCARTHY et al 2015, JOEL 2011, 2012).
Es ist bekannt, dass Lernerfahrungen wie Taxifahren, Tanzen, Jonglieren und sogar Tetris spielen sowohl die Struktur wie auch die Funktionen im Gehirn ändern (MAY 2011, SHORS 2016). WRAGA et al. (2006) demonstrierten, dass bei Durchführung von Untersuchungen zur räumlichen Orientierung sich bei unterschiedlichen Aufgabenstellungen, je nachdem ob man behauptet, dass Männer besser oder schlechter bei solchen Aufgaben sind, sich nicht nur die Testergebnisse, sondern auch die Aktivitätsmuster im Gehirn ändern. Bei anderen Studien wurde auch eine Assoziation zwischen objektiv gemessenem sozioökonomischem Status und Gehirnstruktur gezeigt (HACKMAN & FARAH 2009). Aber auch der wahrgenommene sozio-ökonomische Status, oder wo man sich in einer gesellschaftlichen Hackordnung befindet, kann die Struktur des Gehirns beeinflussen, unabhängig von anderen möglichen Variablen wie ethnischer Zugehörigkeit oder psychischer Gesundheit. (GIANAROS et al. 2007). Es ist mittlerweile auch bekannt, dass Umwelteinflusse auch unser Hormonsystem beeinflussen, die wiederrum auf das Gehirn und unser Verhalten zurückwirken (VAN ANDERS & WATSON 2006, VAN ANDERS 2013). Steroide und andere Hormone gelten dabei als Mediatoren der Verhaltensplastizität, was Tieren ermöglicht auf bestimmte Umwelteinwirkungen kurz- wie längerfristig zu reagieren (OLIVEIRA 2009). Hormone wie Testosteron und Rezeptoren (Moleküle, an die Hormone binden) werden vom Verhalten und sozialer Umgebung moduliert. Beispielsweise reduziert Vaterschaft den Testosteronspiegel (GETTLER et al. 2011). GREENOUGH, BLACK & WALLACE (1987) belegen, dass eine reiche, abwechslungsreiche Umwelt zum Wachstum von Neuronen und Neuer Synapsen führt. MASCIE-TAYLOR (1993) dokumentiert folgende Variablen, die unser Gehirn beeinflussen können: unsere häusliche Umgebung, Komplikationen bei der Geburt, Rauchen während der Schwangerschaft, Beruf, Familie, Schule, geographische Region in der man aufwächst, Kurzsichtigkeit, Ernährung und Anreicherung von Blei. Die Liste von umweltvariablen ist quasi unendlich. Kinder, deren Mütter während der Schwangerschaft geraucht haben, haben z. B. häufig ein geringeres Geburtsgewicht und haben später Probleme bei der räumlichen Orientierung, auch wenn längst nicht alle (BROOKS-GUNN et al. 1992). Musikalisches Lernen hat ebenfalls einen Einfluss darauf, dass sich im Gehirn funktionelle Änderungen entwickeln (HERDENER et al. 2010).
Ein weiteres Anzeichen dafür, dass unser Gehirn sehr plastisch und anpassungsfähig ist, zeigt die Entwicklung unserer synaptischen Verbindungen, also der Kontaktstelle zwischen den Nervenzellen. Denn diese synaptischen Verbindungen werden immer wieder abgebaut, als auch erneuert. Wir lassen dies BECK (2014) kommentieren:
„[m]indestens genauso wichtig [wie der Aufbau neuer Nervenverbindungen – Internet-Evoluzzer] ist der Abbau von Verbindungen,. Erst wenn ein Netzwerk die nötige Plastizität bekommt, die Fähigkeiten sich neuen Reizen und Informationen dynamisch anzupassen, kann es auch neue Dinge lernen. Je mehr Möglichkeiten es gibt, wie sich das Netzwerk verändern und seine Architektur optimieren kann, desto besser und effizienter wird die Informationsverarbeitung sein. (…) Gerade in den ersten Lebensjahren wird von dem Absterben (…) reger Gebrauch gemacht. Es ist übrigens der Grund dafür, weshalb wir uns nicht an unsere Geburt erinnern können: Zu diesem Zeitpunkt ist das Gehirn übermäßig plastisch und wirft Unmengen an überschüssigen Synapsen und Nervenzellen weg, da bleiben die Erinnerungen auf der Strecke. Erst mal muss das Gehirn die grundlegenden Vorschaltungen ausbilden und behält daher lieber Denkmuster (also Verfahren, um überhaupt mal richtig zu denken) als konkrete Erinnerungen.“ (BECK 2014, S. 124)
Merken Sie sich diese Aussage von Beck, wenn sie bei KUTSCHERA (2018, S. 233) lesen, dass die hirnanatomischen und kognitiven Unterschiede bei den Geschlechtern schon bereits im Kleinkindalter festgelegt seien, also biologisch und nicht kulturell determiniert seien.
Zitieren wir weiter BECK (2014, S. 80 – 81):
„Dass Nervenzellen absterben und nicht permanent ersetzt werden, ist unverzichtbar, damit das Gehirn prinzipiell funktioniert! Denn das Allerwichtigste im Gehirn überhaupt ist seine Struktur, die Architektur seiner Vernetzungen. In dieser Vernetzung sind alle Informationen gespeichert, unsere Erinnerungen und Gedanken, genauso wie die Fähigkeit, Sinneswahrnehmung zu verarbeiten oder Bewegungsimpulse zu erzeugen. Dieses Muster der Verknüpfungen muss stabil sein, denn es ist der kostbarste Schatz unseres Denkorgans. Interessanterweise bilden sich die Feinheiten dieser Architektur ohne konkreten Bauplan aus: Sie passen sich im Laufe des Lebens an. Natürlich, die grobe Struktur eines Hippocampus oder des Kleinhirns ist festgelegt, doch entscheidend sind die individuellen Verknüpfungen der Nervenzellen – und diese sind flexibel und dynamisch. (…) Dass die Architektur des Nervenzell-Netzwerks bestimmt, wie das Gehirn funktioniert, hat einen Rahmenvorteil: Es ist robust. Denn es verlässt sich nicht auf die Funktion von einigen wichtigen Nervenzellen (die es natürlich zweifelsfrei gibt, man denke nur an die Bewegungs-Kontroll-Neuronen, die bei Parkinson absterben), sondern auf die Power des ganzen Netzwerks. Das Internet ist ja auch nicht so einfach abzuschalten. Wenn ein wichtiger Server ausfällt, werden die Informationen eben über den nächsten weitergeleitet.„
Am entscheidendsten ist natürlich die Kindheitsentwicklung für die Gehirnentwicklung, da die Anzahl (nicht aber unbedingt die Struktur) der Synapsen vom 2. Bis 10. Lebensjahr relativ konstant bleibt (HALPERN 2012, S. 225). Das heißt natürlich nicht, dass das Gehirn sich im Alter nicht verändert, aber die Dynamik nimmt relativ ab. Übrigens: Mit dem 5. Lebensjahr hat das Gehirn etwa 95% der Masse eines Erwachsenen erreicht, was die großen Kinderköpfe erklärt (HALPERN 2012, S. 225). Mit dem Alter baut sich die Gehirnmasse ab. Mit etwa 70 Jahren reduziert sich das Hirnvolumen um etwa 6% (HALPERN 2012, S. 227). Es zeigte sich, dass die Alterung im Gehirn bei den Geschlechtern unterschiedlich verläuft, wobei Männerhirne schneller altern als weibliche. Dies hat u. a. auch mit den Hormonrezeptoren zu tun, die im Gehirn unterschiedlich verteilt sind. Östrogene scheinen der Alterung entgegenzuwirken (vgl. GUR et a. 1991). Aber auch hier ist eine gewisse Vorsicht geboten: Lebensumstände können einen großen Teil dazu beitragen, wie das Gehirn altert und die Unterschiede im Alterungsprozess bei den Geschlechtern sind minimal (vgl. FJELL, WALHOVD & FENNEMA-NOTESTINE 2009, FJELL et al. 2009).
Weiterhin ist anzumerken, dass die Entwicklung der Genitalien bei Menschen wesentlich früher abgeschlossen ist, als die Gehirnentwicklung. Unser Gehirn ist bei der Geburt ziemlich unterentwickelt (anders würde der großer Kinderkopf nicht durch den Geburtskanal passen).
Nicht nur bei uns Menschen ist das Gehirn anpassungsfähig: Bei einer Studie an Ratten entdeckt man, dass es bei bestimmten Hirnregionen geschlechtsspezifische Unterschiede bei der Form der Dendriten der Nervenzellen gibt, wenn die Tiere stressfrei du friedlich gehalten werden. Dendriten sind jene Bereiche einer Nervenzellen, die die Informationen anderer Nervenzellen aufnehmen. Zwischen beiden Nervenzellen befindet sich der synaptische Spalt, bei dem Neurotransmitter ausgeschüttet werden, sodass die Informationsweiterleitung von einer Zelle zur nächsten übertragen wird. Werden aber die Tiere 15 Minuten unter Stress gesetzt, sehen die Dendriten der männlichen Tiere aus, wie jene der ungestressten Weibchen. Die Dendriten gestresster Weibchen ähneln dann den Dendriten ungestresster Männchen. Stress kehr also die typischen Geschlechtsunterschiede des Gehirns einfach um (SHORS et al. 2001). Es wird aber noch komplizierter. Der selbe Umweltfaktor kann für einige Gehirn-Charakteristika einen Geschlechtsunterschied bewirken, aber den gegenteiligen Einfluss, oder gar keinen, auf andere. Am unteren Ende derselben Hirnregion, die SHORS et al. (2001) untersuchten gibt es unter stressfreien Bedingungen keine Geschlechtsunterschiede bei den Dendriten. Leiden die Tiere aber unter Stress, gibt es bei Weibchen keine Veränderungen, wohl aber bei Männchen. Hier geht es nur um einen simplen Umweltfaktor. Nun stelle man sich vor, dass mehrere tausend solcher Faktoren auf das Gehirn einwirken.
Basierend auf den Erkenntnissen Pawlows und seiner Schüler, wurde in Bezug zur Plastizität der menschlichen Psyche die sogenannte Tätigkeitstheorie ausgearbeitet. Wichtige Vertreter dieser Theorie sind Alexei Nikolajewitsch Leontjew, Lew Semjonowitsch Wygotski und Alexander Romanowitsch Lurija. Außerhalb der Sowjetunion ist diese Theorie auch als „Kulturhistorische Theorie“ bekannt und hat ihre entsprechenden Vertreter, z. B. Jean Piaget. Nach der Tätigkeitstheorie ist das Verhältnis von Mensch und Umwelt ein gesellschaftliches, gekennzeichnet durch die Entwicklung kultureller Werkzeuge und Zeichen. Hauptaussage der Tätigkeitstheorie war, dass nicht die äußere Tätigkeit des Menschen von seinem Bewusstsein abhänge, sondern dass sich das menschliche Bewusstsein andersherum erst durch die gegenständliche Tätigkeit des Menschen in der Welt bilde. Grundannahme der Tätigkeitstheorie ist, dass die menschliche Psyche entsteht und sich stets weiterentwickelt. Sie kann dabei aber nur verstanden werden im Kontext mit bedeutungsvollen, zielorientierten und soziologisch relevanten Interaktionen zwischen Menschen und ihrer materialisierten Umgebung. Aus diesem Grund wurde die Tätigkeit als zentrale Kategorie psychologischer Forschung angenommen. In Zusammenhang mit ihrer psychologischen Arbeit interessierten sich Wygotski, Lurija und Leontjew ab den späten 1920er Jahren zunehmend für Neurologie.
Zusatz-Info:
Es besteht das Gerücht, dass Wygotskis Theorien in der Sowjetunion der 1930er Jahre unter Stalins Führung repressiert wurden, u. a. wegen des 1936 erlassenen Dekrets zur Pädologie, welches u. a. psychologische Tests an Kinder verbat. Jedoch steht in diesem Dekret weder etwas über Wygotskis Theorien, noch seiner Anhänger und das Dekret verurteilt die Auffassung einiger Pädologen, dass sie das Schicks von Kindern aufgrund nicht ändernder biologischer und sozialer Faktoren festlegen. Aber genau das tat die Tätigkeitstheorie von Wygotski, Leontjew und Lurija eben nicht! Es gab sicherlich wissenschaftliche Auseinandersetzungen der verschiedenen Schulen, aber die hat man in jedem Land. Die Pädologie als Wissenschaft, sowie die Lehren von Wygotski wurden aber nicht verboten, denn gerade die Funktion der Sprache und der Arbeit, ein Konzept von Wygotski zur höheren Nerventätigkeit bei Menschen, wurde aktiv erforscht. Zwei Studien belegen dies: FRASIER & YASNITSKY (2015), die zwar zeigen, dass zwischen 1934-1956 die Werke Wygostkis immer seltener publiziert wurden, gleichzeitig aber andere Theoretiker seiner Schule offen praktizierten, sowie eine Publikation der marxistisch-leninistischen Zeitschrift LALKLAR, welche eine Verbindung des Marxismus-Leninismus (inklusive Stalin) mit den Theorien Wygotskis nachweist. Sie erklären ein mögliches Ausbleiben der Publikationen Wygotskis mit dem Aufkommen einiger dogmatischer Richtungen in der Sprachwissenschaft der Schule des Linguisten Marr, welcher von Stalin kritisiert wurde (vgl. Fußnote 19
Es kann ein dieser Stelle kein vertiefter Einblick in die Erkenntnisse und Theorien der Pawlow-Schule und der Tätigkeitstheorie gemacht werden. Interessierten Lesern empfehle ich neben dem LALKLAR-Beitrag zu Wygotski , das Buch „One is not born a personality“ von LEVITIN (2009), „Basic Instinct“ von Mark S. BLUMBERG (2005) und Harry W. WELLS (1989): „Sigmund Freud – Kritik auf Grundlage der Erkenntnisse Iwan P. Pawlows, sowie das Freidenker Heft Nr. 3. 2018 „Menschenbild – Marxistische Persönlichkeitstheorie: Individuum und Gesellschaft“. Natürlich schadet es nicht, Pawlow, Wygotski, Leontjew und Lurija im Original zu lesen.
Festzuhalten ist hier: Aufgrund der Tatsache, dass der Mensch ein soziales Wesen ist, wird sein Gehirn, seine Psyche maßgeblich von der Gesellschaft geformt. Um es aber vorwegzunehmen, da Leute wie CAHILL (2014) vorwerfen, dass solche Theorien über die Plastizität des Gehirns als eine moderne Variante der widerlegten Tabula-rasa-Idee, also der Annahme, das menschliche Gehirn sei wie ein unbeschriebenes Blatt und durch Erfahrungen beliebig veränderbar, sei sofort betont:
Ich bin Gegner des Behaviorismus (zu denen fälschlicherweise Pawlow gezählt wird), des Soziologisierens und der Abtrennung des Menschen von der Natur. Wir sind das Produkt biologischer Evolution und der Arbeit. Gerade die Erkenntnisse von Iwan Pawlow und der Tätigkeitstheorie belegen, dass die angeborenen, unbedingten Reflexe eine wichtige Grundlage für die psychologische Entwicklung des Menschen sind. Gleichzeitig bin ich aber auch ein Gegner des Biologisierens der menschlichen Psyche. Nicht aus politischen Gründen, sondern weil sie wissenschaftlich falsch ist, wie obige Beispiele gezeigt haben und es untere noch tun werden. Unsere Natur ist die Kultur und diese wirkt in einem besonderen Maße auf uns zurück.
Dies entspricht auch den biopsychologischen Hypothesen, die HALPERN (2012, S. 14 ff) vertritt. Die biopsychologische Hypothese basiert auf der Idee, dass einige Unterschiede zwischen den Menschen sowohl biologisch als auch sozial sind und daher nicht in die eine oder andere Richtung klassifiziert werden können. Jedes Verhalten ist – Pawlows System der unbedingten und bedingten Reflexe entsprechend – ein Ergebnis der Interaktion zwischen Genen, Hormonen, Gehirn und Umwelt (FOX, LEVITT & NELSON 2010). Angenommen es bestehe ein natürlicher Unterschied in Bezug zur räumlichen Orientierung zwischen Männern und Frauen aufgrund des Testosteronspiegels. Aufgrund dieses Unterschiedes neigen Männer dazu ihre Fähigkeiten anders zu nutzen als Frauen. Aber die Gesellschaft kann dafür sorgen, dass sich diese zunächst kleinen Unterschiede in gesellschaftlich große verwandeln, da die Interessen für Männer mehr gefördert werden, als für Frauen.
Kurioserweise werden diese Erkenntnisse in den Massenmedien und vielen populärwissenschaftlichen Darstellungen ignoriert. Es wird eine monokausale Kette zwischen Gehirn-Hormonen-Verhalten attestiert, ohne dabei zu berücksichtigen, dass wir hier nicht nur einen Sprung von der Quantität zur Qualität haben, sondern, dass Umwelt, Sozialleben und Lernen das Gehirn ebenso mit formen. Entsprechend haben nur wenige Neurostudien, die die Geschlechterunterschiede zwischen Männer- und Frauengehirnen untersuchten, mögliche Umwelteinflüsse in ihren Ergebnissen berücksichtigt (BISWAL et al. 2010, RIPPON et al. 2014, FINE et al. 2013).
Das Analyse-Problem
Viele Gehirnstudien haben nur sehr wenige Probanden, sodass sich schwer statistische Aussagen machen lassen. Dass die Zahl der Probanden so gering ist, hat auch mit der Aufwendigkeit der Versuche zu tun (fMRTs sind sehr aufwendig, zeit- und kostenintensiv). Abhängig von der Datengröße kann es natürlich zu erheblichen Abweichungen in den Ergebnissen kommen. Hat man eine Studie von 10 Personen, mit z. B. 9 Männer und einer Frau, können die Ergebnisse der gemessenen Unterschiede recht groß ausfallen. Daher kommen auch teilweise so verschiedene Ergebnisse heraus (Probleme der Messmethoden kämen hinzu – leider scheint es bei den meisten Gehirn-Versuchen der Fall zu sein, dass man eben vor der Messung das Geschlecht der Personen weiß, was Einfluss auf das Ergebnis haben könnte). Je größer die Datenmenge aber wird, desto geringer fallen diese Unterschiede auf und verschwinden teilweise ganz. Hier spielen sogenannte Meta-Studien bzw. Meta-Analysen eine wichtige Rolle. Hierbei handelt es sich um eine Zusammenfassung von Primär-Untersuchungen zu Metadaten, die mit quantitativen und statistischen Mitteln arbeitet. Sie versucht frühere Forschungsarbeiten quantitativ bzw. statistisch zusammenzufassen und zu präsentieren. Wenn also diverse Einzelstudien zu einer Forschungsfrage (z. B. Unterschiede im Corpus Callosum) durchgeführt werden, fasst die Meta-Studie diese zusammen, um so ein allgemeineres Bild zu haben. Gerade in Bereichen, die kleine Stichproben haben und/oder große Unterschiede aufzeigen sind solche Metaanalysen zum Verständnis der Forschung unerlässlich. Neben der Vorteilen einer Metaanalyse gibt es natürlich, wie bei jeder statistischen Auswertung, Nachteile. Ein Problem ist, dass Metaanalysen nicht zwischen gut durchgeführten und minderwertigen Studien unterscheiden. Nicht jede Studie ist gut durchgeführt (z. B. zu wenige Probanden, schlechte statistische Auswertungen etc.; vgl. HALPERN 2012, S. 75 ff.). Weiterhin kann es zwar sein, dass Metaanalysen z. B. keine durchschnittliche Unterschiede bei Männern und Frauen finden, es aber z. B. einige „Ausreißer“ gibt: beispielsweise zeigt sich das bei IQ-Tests: durchschnittlich gibt es bei Männern und Frauen keinen Unterschied, wobei der IQ bei Frauen gleichmäßiger verteilt ist, während bei Männern häufiger Extremwerte vorkommen (Idioten & besondere Schlauköpfe). Das heißt, dass sich innerhalb einer Gruppe die Unterschiede stärker sein können, als zwischen zwei Gruppen (vgl. BECK 2014, S. 87).
Es gibt aber ein weiteres Problem: Studien die keine Unterschiede finden werden viel seltener publiziert als Studien, die Unterschiede entdecken können. Angenommen, dass von 100 Studien 90 keine Unterschiede in kognitiven Fähigkeiten finden, so werden diese nicht, bzw. nicht so einfach veröffentlicht. Das hat u. a. damit zu tun, dass wenn Studien Unterschiede finden wollen, es aber nicht tun, für die Publikation in Fachzeitschriften nicht interessant sind und so oft durch das „Peer Review“ (also der Kontrolle durch andere Wissenschaftler, die sich die Publikation näher anschauen und nach Fehlern suchen) nicht kommen oder in kleineren, unbedeutenden Zeitschriften (die vielleicht keinem Peer Review unterliegen) veröffentlicht werden. Wissenschaftler, die aber für ihre Metaanalysen nach Unterschieden zwischen den Geschlechtern suchen, ignorieren diese 90 und suchen nach den 10, die Unterschiede feststellen konnten (vgl. HALPERN 2012, S. 76, FINE 2010, S. 133-134). Die Psychologin Melissa Hines stellt z. B. klar, dass durch die Tatsache, dass es interessanter ist einen Unterschied zwischen den Geschlechtern zu publizieren, als wenn kein Unterschied feststellbar ist (HINES 2004, S. 6). In der Tat wird das Geschlecht in vielen Untersuchungen routinemäßig nebenbei abgefragt, ohne konkrete Hypothese. „Wenn 20 Tests Geschlechterdifferenzen prüfen, wird einer davon sicher ein statistisch signifikantes Ergebnis liefern, auch ohne dass es einen Unterschied zwischen den Gruppen gibt“ (FINE 2010).
Dennoch liefern Meta-Studien durchaus bessere Ergebnisse als nur Einzelstudien. BUTTON et al. (2013) konnten belegen, dass statistische Aussagen von Studien in den Neurowissenschaften sehr gering sind, was zur Überschätzung von Effektgrößen und zur geringen Reproduzierbarkeit der Ergebnisse führt.
Übrigens belegt die persönliche Erfahrung, dass Personen, die unbedingt die Unterschiede in den geistigen Fähigkeiten zwischen Geschlechtern (oder anderen Volksgruppen wie Nationalitäten oder sozialen Klassen) feststellen wollen auf Meta-Studien verzichten und stattdessen selektiv einzelne Studien zitieren. Auch Kutschera zitiert in seinem über 240 Einträge langem Literaturverzeichnis keine einzige Meta-Studie zu Unterschieden im Gehirn oder geistigen Leistungen zwischen den Geschlechtern (verweist lediglich nebenbei auf populärwissenschaftliche Literatur wie PINKER 2008 und MAYER 2015), dafür ist er interessanterweise bei 29 von 240 Einträgen Erstautor im Literaturverzeichnis, wovon aber fast alle über Botanik, allgemeine Evolutionsbiologie und Kreationismus handeln und somit wenig zur „Gender-Debatte“ beitragen.
Desweiteren muss noch ein weiterer Aspekt berücksichtigt werden: Mit Menschen kann man nicht dieselben Experimente wie mit Tieren machen. Über den Einfluss von Testosteron und anderen Hormonen auf die Gehirnentwicklung können wir männliche Ratten kastrieren und feststellen, dass ohne Testosteron sich die Genitalien entwickeln (sie entwickeln keinen Penis). Kastriert man weibliche Ratten bleiben die Geschlechtsorgane weiblich (u. a. durchgeführt von JOST 1970). Man kann Versuchstiere mit Hormonen behandeln, Hirnteile entfernen etc. daher wissen wir so viel wie das Hormon- und Nervensystem funktionieren. Doch solche kontrollierten Experimente können natürlich nicht bei Menschen durchgeführt werden. Wir können solche Experimente bei Menschen natürlich nicht machen, unsere Ergebnisse sind daher immer indirekt. Forschungen an Menschen kann nur indirekt erfolgen, ohne dass wir manipulierend eingreifen. Neben MRTs (die wie gesagt nur indirekte Ergebnisse erhöhter Hirnaktivität zeigen, aber nicht, was wir konkret denken bzw. wie das uns sonst beeinflusst), sind Postmortal-Studien (also Untersuchung an toten Personen), Umfragen, psychologische und ähnliche „harmlose“ Tests oder Langzeitbeobachtungen. Das erfordert also sehr viel Vorsicht Ergebnisse, die man durch Befragungen oder psychologische Tests entsprechend zu interpretieren. Hier kommt aber hinzu, dass sehr viele Variablen mit einbezogen werden müssen, da man auch bei psychologischen Forschungen Menschen nicht komplett isoliert aufwachsen lassen kann (vgl. auch JORDAN-YOUNG 2010).
Wenn man übrigens bei Studien Unterschiede zwischen Männern und Frauen findet, heißt das nicht automatisch, dass man weiß, woher diese Unterschiede kommen und was diese bedeuten. Wir werden sehen, dass unsere Gehirne sehr plastisch sind – also auf Umweltreize reagieren und entsprechend veränderbar sind. Wenn wir also beispielsweise einen Unterschied erkennen: z. B. ist bekannt, dass Männer ein besseres räumliches Vorstellungsvermögen haben als Frauen (übrigens wird dieser Unterschied durch „Lernen“ minimiert), heißt das nicht automatisch, dass wir wissen woher diese Unterschiede (sind die Einfluss bedingt durch die Genetik oder durch die Umwelt?) kommen, ob diese veränderbar sind und ob dahinter überhaupt eine (evolutionäre) Bedeutung steckt. Weiterhin heißen tatsächliche Unterschiede nicht, dass die eine Gruppe besser ist als die andere. Unterschiede sind eben erst mal Unterschiede. Männer haben einen Penis, Frauen eine Vagina. Heißt das damit, dass ein Penis für die sexuelle Fortpflanzung besser ist als eine Vagina? Es leuchtet jedem ein, dass beides notwendig ist für die sexuelle Fortpflanzung bei Menschen, woraus keiner schlechter oder besser ist. Warum sollte es bei anderen tatsächlichen Unterschieden anders sein (vgl. HALPERN 2012, S. 4-5).
Folgerichtig stehen wir vor folgenden Problemen:
* MRTs und andere Methoden des „Hirnscans“ zeigen wertvolle Informationen über die erhöhte Tätigkeit der Gehirne bei bestimmten Aufgaben. Sie haben aber das Problem, dass sie häufig sehr aufwendig sind und die Studienproben entsprechend klein. Daher können sie nicht immer allgemeingültige Aussagen liefern.
* Metaanalysen können hierbei hilfreich sein, doch haben sie zum einen das Problem, dass sie nicht immer zwischen „guten“ und „schlechten“ Studien unterscheiden können. Außerdem: Studien, die keine Unterschiede zwischen den Geschlechtern finden, werden seltener publiziert und man kann diese nicht mit einbeziehen.
* Da wir aus ethischen Gründen nicht am Menschen herum pfuschen können, könnten wir Erkenntnisse über die menschliche Psyche nur indirekt messen: entweder über Tierversuche (die ebenfalls nicht direkt übertragbar sind, siehe nächsten Abschnitt) oder über nicht eingreifende Langzeitbeobachtungen.
* Tierversuche sind aber, trotz Ähnlichkeiten, nicht direkt auf Menschen übertragbar, da es auch, besonders im Gehirn, entscheidende Unterschiede gibt
* Wenn man Unterschiede entdeckt, heißt es noch lange nicht, dass man weiß woher diese Unterschiede kommen. Häufig sind Vorstellungen über die evolutionäre Bedeutung kognitiver Leistungen nur Interpretationen, über die wir gar nicht wissen können, ob sie diese Bedeutung haben. Zumal die evolutionäre Psychologie eben häufig „just-so-stories“, also Anekdoten als gegeben ansehen und unsere heutige (kapitalistische, anglo-amerikanische) Gesellschaft auf die Steinzeit übertragen (vgl. ROSE & ROSE 2001).
* Außerdem heißt es nicht, dass z. B. Frauen schlechter sind als Männer, nur weil man Unterschiede festgestellt hat.
Leider werden besonders in den Massenmedien die Ergebnisse der Neurowissenschaften für eine politische Agenda missbraucht werden. Dabei werden die hier angegebenen Probleme der wissenschaftlichen Methodik völlig ignoriert und daher Unterschiede zwischen Männern und Frauen bezüglich Gehirn und kognitiver Leistungen ignoriert. FINE (2010) prägt den Begriff „Neurosexismus“, um den Missbrauch der Neurowissenschaften zur Rechtfertigung traditioneller Rollenbilder einen Namen zu geben. Mit einem cleveren Wortspiel bezeichnet sie den verantwortungslosen Umgang mit Ergebnissen der Neurobiologie als „brain scam“ (deutsch: Hirnschwindel, abgeleitet von brain scans = Hirn-Scans). Unterstützer der These, dass Männer- und Frauenhirne völlig unterschiedlich sind (z. B. BARON-COHEN 2004, BRIZENDINE 2006, SAX 2005, KUTSCHERA 2018), zitieren gerne tatsächliche und angebliche Unterschiede zwischen Männer und Frauenhirnen ohne zu verstehen, dass der Sprung von Neuronen zum konkreten Verhalten ein sehr großer ist.
Wir werden uns jetzt mit einigen (angeblichen) Unterschieden in Bezug zum Gehirn bei Männern und Frauen befassen.
Hirnrinde
Wir haben schon in unserem historischen Abschnitt festgestellt, dass das Vermessen von Gehirnen eine intensiv durchgeführt Arbeit war. Das ist auch heute nicht anders, vor allem wenn es um den Unterschied zwischen Männer- und Frauenhirne geht. Tatsächlich lassen sich einige Unterschiede ausmachen:
Der präfrontale Cortex ist Teil des Frontallappens der Großhirnrinde (er befindet sich an der Stirnseite des Gehirns) Der präfrontale Cortex empfängt sensorische Signale und steht in korrelativem Zusammenhang mit der Integration von Gedächtnisinhalten und emotionalen Bewertungen. Der ventrale (untere) Bereich des präfrontalen Cortex besteht aus dem orbitofrontalen Cortex. Es befindet sich direkt über der Augenhöhle (Orbita). Dieser Bereich ist evolutionär relativ jung und wurde im Verlauf der menschlichen Evolution größer (u.a. bestätigt durch Schädelausgüssen von Neandertalern, Homo errectus und H. sapiens). Man nimmt an, daß der orbifrontale Cortex eine wichtige Rolle beim Korrigieren der Bewertung emotionaler Stimuli spielt, d.h. wichtig für erlernte Emotionen ist.
Eine Großhirnwindung im Ventralbereich des frontalen Cortext ist der Gyrus rectus, dessen Funktion nicht gänzlich geklärt ist (vgl. Abb. 3).
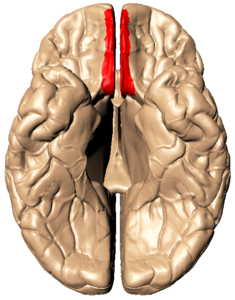
Abb. 3: inferiore/caudale Ansicht des Gehirns, man betrachtet das Gehirn also von seiner unteren Seite. Rot markiert ist der Gyrus rectus
In zwei kleinen Studien zeigte sich dass dieser Bereich des ventralen Frontal-Cortex bei Frauen ein wenig größer ist, was mit der möglichen besseren Fähigkeit von Frauen zusammenhängen könnte soziale Interaktionen einzugehen. Dabei gab es zwischen den Geschlechtern keinen Unterschied im orbitofrontalen Cortex, aber der Gyrus rectus war bei Frauen proportional größer. Die Studie entdeckte auch eine Korrelation zwischen der Größe des Gyrus recuts mit einer größeren Identifikation mit weiblichen Charaktereigenschaften und einem besseren Abschneiden in Tests über soziale Kognitionen (WOOD, HEITMILLER, ANDREASEN & NOPOULOS 2008). In einer weiteren Studie mit 74 Jungen und Mädchen konnte die selbe Arbeitsgruppe jedoch belegen, dass es zwischen Jungen und Mädchen keine Unterschiede im orbitofrontalen Cortex gab, der Gyrus rectus hingegen bei Jungen größer war. Das war das genaue Gegenteil der vorherigen Studie. Die Größe des Gyrus rectus korrellierte dabei negativ mit dem Alter. Das heißt, bei den älteren Jungen war der Gyrus rectus verhältnismäßig kleiner als bei jüngeren, während sich bei Mädchen dieser Effekt nicht zeigte (WOOD, MURKO, NOPOULOS 2008). Dies hängt mit dem Gehirnwachstum zusammen, bei dem die „Graue Substanz“ (der Bereich im Gehirn, der hauptsächlich aus den Nervenzellkörpern besteht, während die „Weiße Substanz“ hauptsächlich aus den Leitungsbahnen – Axonen – der Nervenzellen bestehen) bis ins Teenageralter bei beiden Geschlechtern wächst und danach abbaut. Bei Frauen wird die Graue Substanz im Gyrus rectus jedoch nicht abgebaut, wohl aber bei Männern. Interessant an dieser Studie von WOOD, MURKO, NOPOULOS (2008) ist auch die Tatsache, dass es einen Zusammenhang zwischen dem Gyrus rectus und selbsternannter/interpretierter Weiblichkeit (also weiblichen Charaktereigenschaften) gibt. Bei Erwachsenen fiel die Selbstbeurteilung so aus: assoziiert man sich mit „typisch weiblichen“ Charaktereigenschaften, war der Gyrus rectus größer, bei Kindern hingegen war es genau umgekehrt: weibliche Eigenschaften korrelierten mit einem kleineren Gyrus rectus. Die Forscher kommen zu dem Schluss, dass „die Ursprünge des Zusammenhangs zwischen Sexualdimoprhismus und der Moprhologie des Gyrus rectus und der sozialen Kognition noch nicht aufgeklärt sind.„
Im 19. Jh. hatten Wissenschaftler den Sitz des Intellekts und abstrakten Denkens im Frontallappen behauptet. Untersuchungen an männlichen und weiblichen Hirnen hatten dabei gezeigt, dass die Frontallappen bei Männern größer und komplexer sind, während bei Frauen die Parietallappen (an der Scheitellregion) größer sind. Nachdem aber festgestellt wurde, dass die Parietallappen beim abstrakten Denken stärker beteiligt sind, wurden die Paritellappen auf einmal bei Männern mehr entwickelt (FINE 2010, S. 146).
Tatsächlich können Hirnareale auch ihre Aufgaben wechseln. Eine Studie von BIRD et al. (2004) fand hierzu einige nennenswerte Hinweise. Ein Patient erlitt großflächige Verletzungen des medialen präfrontalen Cortex nach einem Schlaganfall. Das Ausmaß der Schäden war so umfangreich, dass praktisch alle Hirnbereiche, die für das Erkennen von Gedanken anderer (mind reading) notwendig sind, betroffen waren. Interessanterweise war aber der Patient genau dazu in der Lage, konnte also zwischen glücklichen und traurigen Personen unterscheiden. BURACAS, FINE & BOYNTON (2005) fanden heraus, dass die V1 Region (primärer visueller Cortex) bei der Bewegungswahrnehmung stärker aktiviert wird, als die MT-Region (mediotemporaler Cortex, auch V5). Neurophysiologische Untersuchungen bei nichtmenschlichen Primaten ergaben aber, dass die MT-Region stärker aktiviert ist als die V1-Region.
Grundsätzlich ist auch nicht davon auszugehen, dass eine höhere Aktivität automatisch eine höhere psychologische Leistung vorliegt. Die Aktivitätsmuster können z. B. schwächer werden, wenn die Kompetenz der Leistungen steigt und sich weiterentwickeln (BLAKEMORE et al. 2007, BURNETT et al. 2009, HAIER et al. 1992). FRISTON & PRICE (2001, S. 275) bestätigen, dass sich zwar einzelne Hirnareale spezialisieren, diese aber dynamisch und kontextabhängig sind. Beispielsweise kann eine bestimmte Gruppe von Neuronen im temporalen Cortex zu unterschiedlichen Zeitpunkten sowohl die Identität (Wessen Gesicht sehe ich?) wie auch den Gesichtsausdruck (Ist er traurig oder glücklich?) wahrnehmen. Was diese Neuronen machen hängt nämlich von den Informationen ab, die sie von „höher geschalteten“ Neuronen erhalten.
Graue und Weiße Substanz
Man spricht von der Grauen Substanz, wenn man sich auf die Ansammlung von Nervenzellkörpern und ihrer synaptischen Verbindungen bezieht. Die Nervenleitbahnen (Axone) werden als Weiße Substanz bezeichnet. Grundsätzlich wird angenommen, dass Frauen eine dickere Schicht an grauer Substanz haben als Männer (vgl. z. B. LENROOT & GIEDD 2010). Berücksichtig man jedoch die Gesamtgröße des Gehirns, wird das Verhältnis von Weißer zur Grauer Substanz ausgeglichen (LEONARD et al. 2008). Es gibt in Bezug zur Grauen Substanz des Gehirns eher Ähnlichkeiten bei kleineren Gehirnen unabhängig vom Geschlecht (LUDERS, STEINMETZ & JANCKE 2008). Wenn sich also Unterschiede zwischen männlichen und weiblichen Gehirnen finden, sind sie eher ein Effekt der Größe und nicht des Geschlechts. Die meisten Gehirnfunktionen sind das Resultat der verteilten neuronalen Netzwerke und innerhalb dieser findet man eine Komplexität von Verbindungen, Neurotransmittern und synaptischer Funktionen (GIEDD et al. 2006). Folgerichtig spielt der Anteil der Grauen Substanz bei kognitiven Unterschieden zwischen den Geschlechtern keine oder eine untergeordnete Rolle (HALPERN 2012, Z. B. S. 249).
Corpus Callosum
Es gibt jedoch eine andere Hirnstruktur, die über Jahrhunderte lang als der „Heilige Gral“ der Gehirnunterschiede zwischen Männern und Frauen (aber auch zwischen den Rassen) angesehen wird: Das Corpus Callosum, auch bekannt als Hirnbalken (vgl. Abb. 4). Es ist die Verbindung zwischen der rechten und linken Gehirnhälfte und spielt somit beim Informationstransfer im Gehirn eine unheimlich wichtige Rolle. Aber ist es auch ein Merkmal, das zwischen den Geschlechtern unterschiedlich ist?
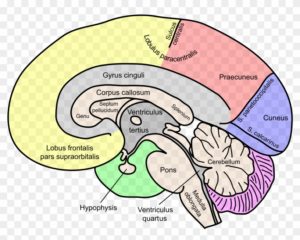
Abb. 4: Längsschnitt durch das Gehirn. In der Mitte der Hirnbalken (Corpus callosum) mit dem vorderen Genu und dem hinteren Splenium
Schon vor über 100 Jahren, genauer 1906, hatte ein Robert Bennett Bean einen längeren Artikel verfasst, der schwarze und weiße US-Amerikaner miteinander verglich. Er hatte einige „sehr auffallende“ Unterschiede entdeckt, besonders beim Corpus Callosum, indem er die Länge des vorderen Abschnittes (Genu, vgl. Abb. 4) des Corpus Callosum mit dem hinteren (Splenium, vgl. Abb. 4) verglich. Dabei stellte er fest, dass die Weißen einen besonders großen Genu hatten, während bei Schwarzen das Splenium länger war. Dieses Ergebnis musste übrigens mit Beans vorherrschender Meinung übereinstimmen: da man die Intelligenz im vorderen Abschnitt des Gehirns vermutete, war es nur erwünscht, dass auch der vordere Teil des Corpus Callosum bei intelligenten Menschen (also den Weißen) logischerweise größer sein musste. Er versäumte auch nicht entsprechende Schlussfolgerungen auch für Frauen zu ziehen: Innerhalb jeder Rasse hätten Frauen ein kleineres Genu als Männer. Er setzte dann seine Abhandlung über die größeren Hirnteile im Stirnbereich bei Weißen fort und schlussfolgerte, dass die Schwarzen ein Zwischenglied zwischen „Mensch (!) und Orang-Utan“ (BEAN 1906, S. 380) seien. Interessant ist nur, dass Bean bei seiner Abhandlung vergaß zu erwähnen, dass Schwarze und Weiße ein gleich großes Gehirn hatten. Doch offensichtlich hatte Bean seine Daten gefälscht. 1909 hatte Beans Mentor Franklin P. Mall Beans Arbeiten wiederholt. Dabei stellte er jedoch sicher, dass er bei der Vermessung der Gehirne nicht erfuhr, ob diese von Weißen oder Schwarzen stammten. Bei einer Stichprobe von 106 Gehirnen konnte er unter Anwendung von Beans Methoden keine Unterschiede im Corpus Callosum feststellen (MALL 1909).
Dieses historische Beispiel sollte uns lehren, dass wenn jemand meint große Unterschiede zwischen den Gruppen gefunden zu haben, man immer sicher gehen sollte, dass der Forscher nicht weiß zu welcher Gruppe die Erforschten gehören.
In den 1990er Jahren tauchten mehrere Publikationen auf, die einen Größenunterschied des Corpus Callosum zwischen Männern und Frauen entdeckt haben sollten. Nun kommt es – man denke an Beans Befunde von 1906: Es wurde gezeigt, dass das Splenium (also der hintere Teil des Corpus Callosum) bei Frauen angeblich größer sei und somit sei geklärt, weshalb Frauen besser ganzheitlicher und integrativer denken als Männer (Bean behauptete übrigens, dass Schwarze aufgrund des größeren Spleniums emotionaler und leidenschaftlicher sind – auch typisch „weibliche“ Attribute).
Nun hatten aber bereits 1997 die Psychologen Katherine Bishop und Douglas Wahlstein eine Metastudie von 49 seit 1980 veröffentlichten Studien der Geschlechtsunterschiede im Corpus Callosum überprüft und konnten keinerlei Unterschiede zwischen den Geschlechtern feststellen (BISHOP & WAHLSTEIN 1997). Auch WALLENTIN (2009) entdeckte keinen Unterschied im Corpus Callosum zwischen Männern und Frauen. Zwar haben einige Analysen in kleinen Bereichen des Corpus Callosum einige Unterschiede feststellen können (z. B. LUDERS, THOMPSON & TOGA 2010), jedoch sind diese Unterschiede zu gering um auch nur daran zu denken, dass sich daraus kognitive Unterschiede zwischen Männern und Frauen ausfindig machen lassen. Die Differenzen dürften maximal bei 0,9% liegen, als dass diese als nennenswerte Variablen anerkannt werden können (für ähnlich geringe Varibalen beim Corpus Callosum oder der Lateralisation siehe auch (BOLES 2005, HAUT & BARCH 2006, LIU et al. 2009). Ein Teil der Komplexität beim Verständnis der Geschlechtsunterschiede im Corpus Callosum besteht darin, dass die Unterschiede in Abhängigkeit vom Alter variieren, weshalb unterschiedliche Studien inkonsistent sind (LENROOT & GIEDD 2010). Außerdem dürfte die Größe des Corpus Callosum auch im Zusammenhang mit der Gesamthirngröße stehen (LEONARD et al. 2008). WELCOME et al. (2009, S. 2434) schlussfolgern: „Es gibt keinen einfachen und universellen Zusammenhang zwischen einer Asymmetrie im Verhalten und der Anatomie des Corpus Callosum„.
Linke und Rechte Gehirnhälfte – die Hemisphären
Wenn aber das Corpus Callosum keinen Unterschied zeigt, dann vielleicht die beiden Hirnhälften? Es wird gerne und häufig spekuliert, dass die verschiedenen Hirnhälften „unterschiedlich denken“. Mathematiker und andere Logiker nutzen die linke Hirnhälfte, weil diese für das logische Denken zuständig ist, während Künstler und andere kreative Menschen eher die rechte Hirnhälfte nutzen (das ist die „empathische“ Hirnhälfte). Es wirkt beinahe so, als ob man zwei Gehirne im Kopf hätte, die unterschiedlicher nicht sein könnten.
Tatsächlich ist unser Gehirn in zwei Teile geteilt, welche über das Corpus Callosum miteinander verbunden sind. Im Grunde genommen ist somit jeder Hirnareal, mit Ausnahme der Epiphyse, in eine linke und rechte Hälfte geteilt. Und sie haben auch unterschiedliche Aufgaben, zumindest teilweise. Tatsächlich passen sich auch beide Hirnhälften im Laufe des Lebens an und verändern an einigen Stellen ihre Form. Bei Rechtshändern liegen 96% des Sprachzentraums auf der linken Hälfte, entsprechend ist diese ein wenig größer. Noch interessanter wird es, wenn man bedenkt, dass wir mit den Hirnhälften die jeweils gegenüberliegende Köperseite steuern. Die rechte Hirnhälfte kontrolliert den linken Arm und anders rum. Es ist aber nicht anzunehmen, dass die beiden Hirnhälften völlig unabhängig voneinander arbeiten – für die Verbindung sorgt das Corpus Callosum. Ein Abtrennen der Gehirnhälften kann dazu führen, dass die Kommunikation zwischen beiden Hirnhälften fehlerhaft ist. Eine Trennung der Hirnhälften ist aber manchmal medizinisch sinnvoll – z. B. bei Epileptikern, eine Überaktivität bestimmten Hirnregionen, die sich dann auf das ganze Gehirn ausbreitet. Eine Trennung des Corpus Callosum verhindert, dass sich solch eine Überaktivität über das ganze Gehirn ausbreitet. Interessant ist hierbei nicht nur, dass die epileptischen Anfälle sich bessern, gleichzeitig keine großen Einbußen der Hirnleistungen zu bemerken sind. Untersuchungen an solchen „split-brain-Patienten“ haben jedoch ergeben, die linke und rechte Gehirnhälfte zum Teil unterschiedliche Funktionen haben. Präsentiert man einen Gegenstand so, dass er im rechten Gesichtsfeld erscheint, dann wird er von der linken Gehirnhälfte erkannt. Da bei den meisten Menschen das Sprachzentrum Großteils in der linken Hirnhälfte zu finden ist, kann der split-brain-Patient den Gegenstand richtig benennen. Lässt man den Gegenstand aber im linken Gesichtsfeld erscheinen, wird das Bild dieses Gegenstandes in der rechten Gehirnhälfte verarbeitet. Da jedoch eine Verbindung zum Sprachzentrum fehlt, kann der Gegenstand nicht benannt werden. Diese Versuchsanordnungen zeigten, dass unsere Hirnhälften tatsächlich Informationen unterschiedliche verarbeiten (BECK 2014, S. 51 – 54). Dies führt aber dazu, dass pseudowissenschaftliche Kommentatoren daraus schließen man hätte dadurch unterschiedliche Eigenschaften, Charaktere – oder gar dass die linke Hirnhälfte die „logische“ und die rechte die „künstlerische“ sei – und je nachdem welche Hirnhälfte dominiert, ist man entweder Logiker oder Künstler … korreliert das mit dem Geschlecht ist der Mythos perfekt. Neurobiologisch macht er jedoch keinen Sinn, trifft nicht zu. Wichtiger Tipp: wann immer man Ratgeber-Bücher, Bildchen etc. sieht, die einem suggestieren wollen, man sei entweder ein logischer oder kreativer Denker, weil unsere Hirnhälften so funktionieren, kann man sich sicher sein, dass diese Aussagen so wissenschaftlich sind, wie die Vorstellung Gott habe die Welt in sechs Tagen erschaffen.
Neurowissenschaftler haben beispielsweise mithilfe von fMRTs untersucht, wenn man kreativ ist und sich vorstellt, improvisierend durch den Raum zu tanzen (FINK et al. 2009). Sie fanden dabei tatsächlich einen Unterschied: Die Hirnaktivität der Probanden zeigt, dass bei einem klassischen Walzer das Gehirn weniger großflächiger aktiviert wird als beim „freien Ausdruckstanz“. Hinzukommt: beide Hirnhälften sind gleichermaßen am Improvisieren freier Bewegungen beteiligt sind. Dies trifft übrigens auf alle möglichen Varianten „kreativen Denkens“ zu, denn je nach Aufgabe sind unterschiedliche Bereiche des Gehirns aktiv. Es gibt keine Region im Gehirn, die verantwortlich ist für die Kreativität (vgl. DIETRICH & KANSO 2010). Das trifft auch für das mathematische, also logische Denken zu. fMRT-Untersuchungen konnten feststellen, dass Matheaufgaben besonders gut zu lösen sind, wenn die beiden Hirnhälften miteinander kommunizieren (SINGH & BOYLE 2004). Das trifft auch auf die Sprache zu: zwar sitzt das Sprachzentrum überwiegend auf der linken Seite. Doch mit der rechten Hirnhälfte geben wir den Wörtern eine Sprachmelodie, also auch hier arbeiten die Hirnhälften zusammen (BECK 2014, S. 56).
Die Annahme, dass Menschen überwiegend entweder mit der linken oder rechten Hirnhälfte denken ist falsch. Untersucht man bei tausend Probanden, wo im Gehirn sie bestimmte Aufgaben verarbeiten, stellt man fest, dass es nur wenige Netzwerke gibt, die auf einer Seite des Gehirns besonders stark ausgeprägt sind. Die meisten Aufgaben werden von verschiedenen Hirnregionen, sowohl in der rechten wie in der linken, verarbeitet (O’BOYLE 2005, O’BOYLE et al. 2005, SINGH & O’BOYLE 2004 NIELSEN et al. 2013). Unsere zwei Hirnhälften bilden ein gemeinsames Netzwerk.
Eine Studie von INGALHALIKAR et al. (2014) ergab, dass die beiden Großhirnhälften von Männern und Frauen unterschiedlich vernetzt sind. Dabei wurden 949 Menschen (428 männlich, 521 weiblich) im Alter von 8 bis 22 Jahren unter Einsatz der Diffusion Tensor Imaging (DTI)-Technik analysiert. Hierbei wurden die neurologischen Verbindungen zwischen den Hirnarealen und den beiden Hirnhälften analysiert. Die Forschergruppe konnte folgende Ergebnisse verzeichnen: bis zum 13. Lebensjahr gab es zwischen den Geschlechtern keine Unterschiede. Mit dem Einsetzen der Pubertät waren jedoch die Gehirne weiblicher Probanden stärker vernetzt, vor allem die Vernetzung der linken und rechten Gehirnhälfte waren deutlich stärker ausgeprägt (vgl. Abb. 5). Bei Verhaltensstudien, die die Forscher ebenso durchführten, zeigte sich, dass Frauen aufmerksamer waren als Männer und sich leichter Wörter und Gesichter merken können. Männer hingegen waren wären bei räumlicher und motorischer Orientierung den weiblichen Probanden überlegen. Die größten Unterschiede zeigten sich dabei bei 12 – 14 jährigen Probanden.
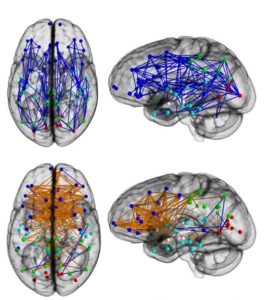
Abb. 5: Vernetzung der Gehirnhälften nach INGALHALIKAR et al. (2014). Oben männlich unten weiblich. Die Ergebnisse zeigen, dass die Hirnhälften bei Frauen stärker vernetzt sind, während bei Männern die Vernetzungen innerhalb einer Hirnhälfte stärker ist.
Die Studie sorgte für Aufsehen, da nun ein weiterer Beweis dafür gebracht wurde, dass Männer und Frauen unterschiedlich denken. Auch KUTSCHERA (2018, S. 231 – 232) zitiert voller Stolz diese Studie. Er untermauert dies auch durch eine positive Bewertung von INGALHALIKAR et al. (2014) durch CAHILL (2014). Es hagelte aber auch an Kritik, die Kutschera interessanterweise verschweigt (dabei hätte er sie doch wenigstens als ehrlicher Forscher erwähnen und kritisieren können?).
JOEL & TARRASCH (2014) kritisierten an der Studie von INGALHALIKAR et al. (2014), dass deren Abbildungen nicht die Realität darstellen, da sowohl männliche wie auch weibliche Gehirne intra- und interhemisphäre Verbindungen haben. Das heißt beide Geschlechter verfügen sowohl über Verbindungen zwischen den Gehirnhälften, wie auch innerhalb einer Gehirnhälfte. Das wäre durch die Darstellungen von INGALHALIKAR et al. (2014) verfälscht, da sie nur die Unterschiede darstellen. Hinzu käme, dass die Autoren nur in einigen dieser Verbindungen Unterschiede zwischen den Geschlechtern fanden. Aber nicht alle Verbindungen der Hirnregionen oder Hirnhälften zeigen Geschlechtsunterschiede. Weiterhin geben die Autoren auch nicht die Größe dieser Unterschiede an, genauso wenig wie stark die Übergänge sind. Dadurch lasse sich aber nicht klären, wie bedeutungsvoll diese gefundenen Unterschiede sind. INGALHALIKAR et al. (2014a) geben eine Antwort auf die Kritik von JOEL & TARRASCH (2014) und merken an, dass die dargestellten Unterschiede die signifikanten durchschnittlichen Unterschiede zwischen den Geschlechtern darstellen. Das wesentliche Ziel ihrer Studie sei es neuropsychiatrische Erkrankungen besser zu verstehen. Keineswegs sind INGALHALIKAR et al. (2014 & 2014a) der Meinung, dass Mann und Frau dadurch komplett unterschiedlich in ihren Denkleistungen sind.
CORNEL (2014) lobt das Manuskript von INGALHALIKAR et al. (2014) und findet viele der Argumente und Darstellungen überzeugend. Sie hat aber auch eine Reihe an ernstzunehmenden Kritiken. Beispielsweise zeigen ihre Daten die Entwicklungsunterschiede der Gehirnverbindungen bei Kindern, Jugendlichen und jungen Erwachsenen an, sie werden aber als generelle Geschlechterunterschiede dargestellt und nicht als sich graduell verändernde Unterschiede reifender Gehirne. Das Fehlen von Geschlechtsunterschieden bei bestimmten Jahrgängen wird zwar im Diskussionsteil, nicht jedoch im Ergebnisteil beschrieben. Zwar geben die Autoren das Alter, Rasse und das Geschlecht der fast 1000 Probanden an, jedoch nicht weitere Differenzen, wie z. B. den Bildungshintergrund und wie man an diese Probanden gekommen ist. Aber auch das hat sicherlich Einfluss auf die Ergebnisse.
RIPPON (2016) kritisiert, dass die „umfassenden“ Ergebnisse von INGALHALIKAR et al. (2014) eher gering waren und sie keine große Effektstärke hatten, statistisch also wenig signifikant sind. Der d-Wert zur Effektgröße bei der Vernetzung der Gehirnhälten ergab einen Wert von 0.309 (RIDGWAY 2013). Der d-Wert nach COHEN (1988) ist die Effektgröße für Mittelwertunterschiede zwischen zwei Gruppen. Cohen’s d wird verwendet um den Effekt über verschiedene Studien hinweg zu vergleichen, selbst wenn die abhängige Variable auf unterschiedliche Weise gemessen wurde. Wer sich für Statistik interessiert, kann über Cohen’s d mehr hier erfahren
Entscheidend ist: ist der d-Wert unter 0.2 ist der Effekt klein, ein d-Wert von 0.5 hat einen mittleren Effekt, ein d-Wert ab 0.8 hat einen großen. Der von RIDGWAY (2013) berechnete d-Wert für die Vernetzung beider Hirnareale aus INGALHALIKAR et al. (2014) Studie beträgt 0.309 und ist damit eher gering (der höchste gemessene Wert der Studie war übrigens 0.482). als Vergleich: der d-Wert für die Körpergröße bei Mann und Frau beträgt nach RIDGWAY (2013) 1.72.
Eine Studie von SATTERTHWAITE et al (2015) untersuchten ebenfalls die Vernetzung der beiden Gehirnhälften. Sie verwendeten jedoch eine andere Methode, nämlich die Resting State fMRT, um die Muster der funktionellen Konnektivität aufzudecken. Die Patienten lagen im Scanner und machten nichts weiter als auf ein Kreuz in einem Bildschirm zu gucken. Hieraus wurde eine Arte „Karte“ hergeleitet, die zeigt, welche Hirnregionen miteinander kommunizieren. 674 Patienten im Alter von 9 bis 22 nahmen daran teil. Dabei mussten sie auch bestimmte Aufgaben lösen: räumliche und motorische Aufgaben, bei dem die männlichen Patienten besser abschnitten, weibliche Patienten waren dagegen beim Erkennen von Emotionen und nonverbaler Kommunikation besser. Darüber hinaus deckten die Forscher auch Geschlechtsunterschiede bei der funktionellen Gehirnverbindung im Ruhezustand auf. Die Unterschiede waren in allen Altersgruppen vorhanden (wobei bei den Unterschieden zu fragen ist, ob diese genetisch oder kulturell bedingt sind – man darf auch nicht vergessen, dass auch Kinder von 9 Jahren eben eine 9-jährige Sozialisation hinter sich haben). Am wichtigsten ist jedoch, dass die Forscher eine bescheidene Verbindung zwischen den beiden Geschlechtern gefunden haben. Je stereotyper das weibliche Gehirn einer Person im Vergleich zu einem männlichen war, desto stärker war ihr kognitives Profil auch stereotypisch weiblich und umgekehrt. Es gab also funktionelle Unterschiede zwischen beiden Geschlechtern im Gehirn, doch diese waren sehr bescheiden:
Um die funktionellen Verbindungsmuster zu untersuchen, untersuchten die Forscher 264 funktionale „Knotenpunkte“ oder Hubs (d. h. Orte im Gehirn, die wahrscheinlich unterschiedliche mentale Aktivitäten unterstützen) und 36.716 spezifische funktionale Verbindungen (d.h. Berührungspunkte zwischen zwei Informationskanäle). Nur sechs der Knoten (2,3 Prozent) wiesen einen Unterschied zwischen den Geschlechtern auf:
Fünf zeigten Anzeichen von mehr Verbindungen innerhalb eines Knotens als mit anderen Knoten bei Männern im Vergleich zu Frauen; der sechste Knoten zeigte bei Frauen eine stärkere Konnektivität innerhalb der Knoten als bei Männern auf. Bei den tausenden untersuchten Verbindungspunkten zeigten nur 0,51 Prozent einen Unterschied zwischen den Geschlechtern. Kurz gesagt, die geschlechtsspezifischen Unterschiede im Gehirn waren winzig. Gegen Ende ihrer Arbeit bestätigten die Forscher diesen Punkt: „Obwohl es geschlechtsspezifische Unterschiede in der Konnektivität gibt“, sagten sie, „sind die Konnektivitätsmuster von männlichen und weiblichen Gehirnen mehr gleich als verschieden.“
INGALHALIKAR et al. (2014) behaupten, aus strukturellen Unterschieden geistige Unterschiede abzuleiten, was aber z. B. durch mehrere Studien eben nicht belegt wird. KAISER et al. (2009) bemerken, dass selbst wenn strukturelle und geistige Unterschiede zwischen Mann und Frau gefunden wurden, sie dennoch keinerlei Auskunft darüber geben, ob diese Unterschiede angeboren sind oder durch „Erfahrung“ oder „Erziehung“ zustande kommen bzw. ob diese Unterschiede auch veränderbar sind.
GESCHWIND & GALABURDA (1985) sowie GESCHWIND & BEHAN (1982) hatten postuliert, dass der höhere Testosteron-Gehalt im Mutterleib während der Schwangerschaft dafür sorgt, dass bei männlichen Individuen sich die linke Gehirnhälfte langsamer entwickele. Dadurch seien Männer potentiell talentierter, was die Fähigkeiten betrifft, die typisch für die rechte Gehirnhälfte sind (Kunst, Musik, Mathe, vgl. z. B. KOLATA 1983 – auf einmal ist ein typisch logisch-linkes Verhalten wie Mathematik Teil der rechten Gehirnhälfte, soso). Diese Hypothese wurde aber von GILLMORE et al. (2007) widerlegt, indem sie das Gehirn von 72 Neugeborenen scannten und keinen Unterschied zwischen den Gehirnhälften entdeckten. Eine Meta-Analyse von SOMMER et al. (2004) mit 377 untersuchten Männern und 442 Frauen fand auch keinen Unterschied in der Lateralisation der Gehirnhälften bei der Sprache (d. h. beide Gehirnhälften waren beim Sprechen beteiligt). Aber schon 1986, als die Theorien von Geschwind „modern“ waren, hatte BLEIER (1986), dass diese Theorie unschlüssig ist, indem sie bei einer groß angelegten postmortalen Studie fötaler Hirne keine Unterschiede zwischen linker und rechter Gehirnhälfte entdeckte.
Eine weitere Meta-Analyse untersuchte den Einfluss des pränatalen Testosteron bei Menschen und anderen Säugetierarten, sowie bei Vögeln auf die Lateralisation des Gehirns. Dabei entdeckten sie, dass pränatales Testosteron einen Einfluss bei nichtmenschlichen Säugetieren (hauptsächlich motorische Eigenschaften) und Vögeln (visuell-kognitive Aufgaben), jedoch nicht bei Menschen (PFANNKUCHE, BOUMA & GROOTHUIS 2008).
KAISER et al. (2009) veröffentlichten eine Bildgebungsstudie, in der bei 44 Probanden das für die Sprachproduktion zuständige Broca-Areal untersucht wurde. Anders als in vorangegangenen Untersuchungen waren bei den männlichen Teilnehmern die Sprachzentren in beiden Hirnhälften gleichermaßen aktiviert, bei den Frauen hingegen dominierte das Broca-Areal der linken Hemisphäre – so schien es zumindest. Doch als die Forscher einfach nur ihre Auswertungsmethode änderten und strengere statistische Maßstäbe anlegten, verschwand der Unterschied auf einmal! Nun war bei beiden Geschlechtern eine stärkere Aktivierung des linken Broca-Zentrums gegenüber dem in der rechten Hirnhälfte zu verzeichnen.
Aber schon in den 1980ern konnte MCGLONE (1980), der die Studien zur zerebralen Asymmetrie erfasste zusammenfassen, dass man die eigentliche Schlussfolgerung nicht außer Acht lassen sollte, nämlich dass die grundlegenden Muster der Hirn-Asymmetrie bei Männern und Frauen im wesentlichen eher gleich sind als unterschiedlich.
Eine Studie von CAHILL (2003) zeigte bei bildgebenden Verfahren, dass bei Frauen, die emotionale Erinnerungen hatten, die linke Hirnhälfte stärker aktiviert war als die rechte. Bei Männern war es anders rum. Schaut man sich aber die Daten näher an, so erkennt man eine große Überlappung bei den Geschlechtern. LIPPA (2005) schlug vor, dass bei Männern die rechte Hemisphäre visuelle und räumliche Informationen verarbeitet, während die linke Hemisphäre hauptsächlich das Sprachzentrum darstelle. Bei Frauen wären die Aufgaben eher verteilt im Gehirn verarbeitet. Diese Aussagen werden aber von der Metaanalyse von SOMMERS et al. (2008) widerlegt (siehe oben). Eine Metaanalyse bezüglich räumlicher Orientierungen fand auch keine starken Geschlechterunterschiede (VOGEL, BOWERS & VOGEL 2003)
FINE (2014) gibt in einem Interview bekannt, dass die Unterschiede zwischen den „Verdrahtungen“ männlicher und weiblicher Hirne auch eine Frage der Gehirngröße ist. Das heißt es spielen konstruktionsmorphologische Aspekte eine wichtige Rolle. Männliche Gehirne sind im Durchschnitt 8-10% größer als weibliche, was Auswirkungen auf die Verbindungen einzelner Hirnregionen haben könnte.
Diese Erklärung erscheint logisch, denn wenn männliche Gehirne größer sind (und 10% mehr Masse ist nicht wenig!), sind Verbindungen zwischen den Gehirnhälften unökonomisch, da diese viel länger sein müssten. Kleinere Frauenhirne können sich diese Vernetzungen eher leisten. Solch ein Zusammenhang zwischen Gehirngröße und Konnektivität konnten auch JÄNKE et al (2014) feststellen. Daraus ist aber auf keine Höherwertigkeit männlicher Gehirne zu schließen bzw. geistige Unterschiede abzuleiten. Die Analyse von CAROTHERS & REIS (2013) demonstrieren gerade diese Befunde. Sie finden zwar typische Geschlechtsunterschiede bei stereotypischen Aktivitäten (Golfspielen, Tragen von Make-up). Die meisten der Unterschiede, die sie entdeckten waren jedoch nicht signifikant und zu der Schlussfolgerung kommen, dass sich zwar Geschlechter in manchen Aktivitäten unterscheiden, diese aber nicht erklären, dass „Männer so sind“ und Frauen anders. Die Unterschiede sind eher als individuelle Unterschiede anzuerkennen, als typische Unterschiede zwischen den Geschlechtern (REIS & CAROTHERS 2014).
Große Hirne sind nicht automatisch besser, da eine Informationsübertragung durch Nervenbahnen länger dauert und energieaufwendiger ist (BECK 2014, S. 67, vgl. auch IM et al. 2008). Folgerichtig sind weniger stark ausgeprägte lange Verbindungen zwischen den Hirnhälften unökonomisch. Tatsächlich finden Studien eher einen Zusammenhang zwischen kleineren Gehirnen und der Feinstruktur unabhängig vom Geschlecht (IM et al. 2008, LEONARD et al. 2008).
BECK (2014) kommentiert:
„Hier sieht man schon das ganze Missverständnis des Rechts/links-Mythos: Die Leute übertreiben und sie bringen Sachen durcheinander. Nur weil konkrete Funktionen im Gehirn vornehmlich auf einer Seite liegen, heißt das noch nicht, dass man damit ganze Charaktereigenschaften erklären kann.“ (BECK 2014, S. 56)
Männerhirne = Systematik / Frauenhirne = Empathie?
Wir haben oben festgestellt, dass Männer ein etwas größeres Gehirn haben als Frauen. Daraus lassen sich aber keineswegs intellektuelle Fähigkeiten ableiten. Jedoch sind längst nicht alle Forscher so vorsichtig. Simon Baron-Cohen ist Autismusforscher und vertretet die sogenannte „extreme male brain theory“. Sie besagt, dass Autisten, verursacht durch einen hohen Testosteronspiegel im Mutterleib, ein extrem ausgeprägt männliches Gehirn haben (BARON-COHEN 2004). Da Autisten für ihre systematische Denkweise und ihren Mangel an Empathie bekannt sind, werden Frauenhirne als „Empathizers“ (Mitfühlenden) und Männerhirne als „Systematizer“ (Sytematiker) dargestellt. Da Männer systematischer denken, sind sie entsprechend besser in logischen Bereichen wie Mathematik, während Frauen eher mitfühlend sind, weswegen sie besser mit Kindern umgehen können.
Cohens Hypothese stieß auf heftige Kritik (z. B. ELIOT 2009, SPELKE & GRACE 2007). Zum Beispiel sind Männer, die noch im Mutterleib höheren Testosterongehalt ausgesetzt waren (sogenanntes CAH-Syndrom), nicht männlicher, als jene mit durchschnittlichen Testosteronwerten. Auch die Theorie, dass ein hoher Testosterongehalt Autismus verursacht, ist nicht bestätigt worden (HALPERN 2012, S. 243). Auch sind autistische Jungen nicht „übermännlich“ (ELIOT 2009).
Man hat damit nicht ein typisches Männer-Gehirn oder Frauen-Gehirn, sondern ein Mosaik aus verschiedenen Eigenschaften (JOEL et al. 2015). Diese Befunde fanden JOEL et al. (2015) als sie Bilder von 1400 menschliche Gehirne aus großen Datenbanken von vier unterschiedlichen Quellen analysierten. Zuerst identifizierten sie die 10 größten Geschlechtsunterschiede in jeder dieser Datenbanken. Aber keiner dieser Unterschiede war wirklich nennenswert, da selbst bei den größten Unterschieden es große Überschneidungen gab. Die Unterschiede waren eher durch das Alter, Umwelt oder genetische Variationen bestimmt als durch das Geschlecht. Außerdem hatte jeder der Datensätze seine eigenen Top-10-Liste. Insgesamt zeigten jedoch nur 23 bis 53 Prozent aller untersuchten Gehirne morphologische Merkmale, die sich tendenziell einem Geschlecht zuordnen ließen. Unter Einbeziehung mehrerer Strukturen zeigten zwischen null und acht Prozent der untersuchten Probanden ein typisch männliches oder weibliches Gehirn. Zwar gibt es Geschlechter-Unterschiede im Gehirn, sie können aber nicht wie die Geschlechtsorgane in typisch männlich und typisch weiblich eingeteilt werden. Viel mehr zeige sich ein Mosaik aus verschiedenen Eigenschaften (JOEL 2011, JOEL et al. 2015). Hinzu kommt, dass Geschlechtsorgane eine entscheidende Funktion haben (Keimzellenproduktion und -verbreitung), das treffe aber auf Gehirne nicht zu. Viele Funktionen der geschlechtsspezifischen Unterschiede im Gehirn sind noch unbekannt und tatsächlich kann nur ein Bruchteil der bekannten Unterschiede im Gehirn zwischen den Geschlechtern eine Verbindung zu geschlechtsspezifischen Verhaltensweisen aufzeigen (DE VRIES & FORGER 2015, DE VRIES & SÖDERSTEN 2009). Beispielsweise können Geschlechtsunterschiede im Gehirn tatsächlich auch potentielle Unterschiede im Verhalten kompensieren (DE VRIES & SÖDERSTEN 2009). Von Ratten weiß man z. B. dass eine kleiner Anzahl von Nervenzellen in einer bestimmten Gehirnregion durch die Produktion von mehr Neurotransmittern kompensiert werden kann (MOORE 1995, S. 95). Dass Gehirnunterschiede zu gleichen Verhaltensweisen führen, hat man u. a. auch bei Prärie-Wühlmäusen entdeckt, bei dem Männchen wie Weibchen bei der Jungenaufzucht gleichmäßig beteiligt sind. Die mütterliche Fürsorge wird bei Prärie-Wühlmaus Weibchen natürlich über das Hormonsystem geregelt. Aber wie entwickeln sich die Vatergefühle dieser Art? Die Antwort liegt in einem bestimmten Gehirnbereich, dem lateralen Septum, der bei beiden Geschlechtern dieser Art unterschiedlich ist. Bei Männchen ist dieser Bereich stärker mit Rezeptoren für das Hormon Vasopressin ausgestattet. Aber genau dieser Unterschied sorgt dafür, dass sich Männchen und Weibchen gleich um die Jungtiere kümmern (MOORE 1995, S. 53 – 54). HAIER et al. (2005) gehen auch davon aus, dass unterschiedliche Gehirndesigns dieselben Aufgaben lösen können. Bei Untersuchungen des Gehirn konnten sie feststellen, dass Männer und Frauen sich zwar in gewissen Regionen im Gehirn unterscheiden, bei Intelligenztest jedoch durch die Aktivität unterschiedlicher Regionen im Gehirn ähnlich abschneiden. Folgerichtig können unterschiedliche Hirnstrukturen zu selben Intelligenzleistungen befähigt sein.
Kritiker an JOEL et al. (2015) Studie bemerken, dass man durchaus statistische Techniken verwenden kann, die aufzeigen, dass es eindeutig männliche und weibliche Hirne geben kann (DEL GUIDICE et al. 2016). JOEL et al. (2016) antworten darauf, dass es keine biologisch bedeutenden Sinn mache anzugeben, welche Gehirne statistisch ähnlicher sind welche nicht. Außerdem zeigen statistische Techniken, dass wenn in einem Datensatz ein erfolgreicher Unterschied festgestellt werden kann, dieser bei einem anderen Datensatz fehlschlägt.
RITCHIE et al. (2018) untersuchten 2750 Frauenhirne und 2466 Männer Hirne, alle im Alter von 44 bis 77 Jahren, nach Unterschieden in den Geschlechtern. Sie untersuchten dabei das Volumen von 68 Regionen im Gehirn und die Dicke der Hirnrinde. Sie fanden heraus, dass (unter Einbeziehung des Alters) die Hirnrinde bei Frauen dicker ist, Männer dafür das größere Gehirn haben, inklusive einzelner Hirnareale, wie Hippocampus und Amygdala, die bei Männern größer waren. Stellt man jedoch die einzelnen Hirnregionen in Relation zur gesamten Hirngröße, wurde der Unterschied kleiner: es gab 14 Regionen, die bei Männern verhältnismäßig größer waren und 10 bei Frauen. Das Volumen und die Dicke der Hirnrinde variierte bei Männern stärker als bei Frauen. Dies passe laut RITCHIE et al. (2018) auch zu den Intelligenztest, bei denen es bei Männern eine größere Variabilität gäbe (in wieweit Intelligentestes aussagekräftig sind, sei erst mal dahingestellt). Neben den Unterschieden stellten sie aber noch etwas fest: eine enorm große Überlappung zwischen Männern und Frauen in Bezug auf das Gehirnvolumen und die Dicke der Hirnrinde. Die Studie gibt – berechtigterweise – aber keine Auskunft darüber, was diese Unterschiede und Gemeinsamkeiten in der Hirnanatomie der Geschlechter für Auswirkungen auf geistige Fähigkeiten haben. Auch RUIGROK et al. (2014) konnten belegen, dass in der Gehirngröße, der Amygdala und im Hippocampus durchschnittliche Unterschiede zwischen den Geschlechtern festzustellen sind.
Die Neurowissenschaftlerin Lise ELLIOT (2009) fand heraus, dass sich die Gehirne von Jungen und Mädchen nur wenig unterscheiden. Die einzigen Unterschiede, die sie entdeckt, wenn sie ALLE Daten berücksichtigt ist, dass die Gehirne von Jungen größer sind, als die von Mädchen und dass Gehirne von Mädchen früher aufhören zu wachsen.
Natürlich gibt es Unterschiede zwischen weiblichen und männlichen Gehirnen, wobei nicht ganz klar ist, welche denn wirklich relevant sind. Zwar sind männliche Gehirne tatsächlich durchschnittlich 8 – 10% größer als die weiblichen. Doch das spielt definitiv keine Rolle. KAISER et al. (2009) betonen, dass die Ergebnisse, die Geschlechtsdifferenzen, anatomisch wie funktionell, im Bereich der linken und rechten Hemisphäre, der Größe des Corpus Callosum und des Ausmaßes definierter Hirnregionen gefunden worden seien, niemals sowohl überzeugend oder unangefochten waren. Der Punkt ist aber derjenige: weder anatomische Details des Gehirns, noch die Darstellungen ihrer Funktionen im fMRT erlauben es, die Unterschiede bei Männern und Frauen zu zeigen. Wenn Unterschiede auffallen, sind diese eher individuell und nicht auf das Geschlecht bezogen.
„Unsere Nachricht zum Mitnehmen heißt, dass es Geschlechtsunterschiede im Gehirn gibt, aber die Gehirne von Männern und Frauen sind einander viel ähnlicher, als dass sie sich unterscheiden.“ (HALPERN 2012, S. 241)
Anhand der Analysen von JOEL et al. (2015), RITCHIE et al. (2018), KAISER et al. (2009) und RUIGROK et al. (2014), inklusive der in den anderen Abschnitten zitierten Studien, lässt sich festmachen, dass sich zwar in einigen Bereichen des Gehirns zwischen Männer und Frauen Unterschiede finden lassen, diese sich aber auf einzelne Bereiche konzentrieren, es große Überschneidungen bei beiden Geschlechtern gibt und die morphologischen Unterschiede alleine keine funktionellen oder kognitiven erklären.
CAHILL (2014a) hält aber weiterhin das Gehirn von Männern und das von Frauen für grundlegend verschieden. Zwar gebe es häufig starke Überlappungen zwischen vermeintlich männlichen und weiblichen Eigenarten, und die gefundenen Effekte fielen häufig bescheiden aus. Doch das liege vor allem daran, dass die Analysen nur isolierte Funktionen betrachten. „Das ist, als würde man nach detaillierter Prüfung von Glas, Reifen und Bremsen zu dem Schluss kommen, dass es keine bedeutsamen Unterschiede zwischen einem Volvo und einer Corvette gibt“. Jene, die die wesentliche Gleichheit der Gehirne bei Männern und Frauen entdecken wollen, wirft er vor eine „anti-Geschlecht-Haltung“ zu haben, die keinen Unterschied zwischen Männern und Frauen sehen wollen. Aber genau das wird von jenen Forschern (JOEL et al. 2015, FINE 2010, JORDAN-YOUNG 2010 etc.) eben nicht behauptet. Sie erkennen Unterschiede zwischen Geschlechtern im Gehirn an, sagen aber, dass diese Unterschiede nicht bedeuten, dass es sich quasi um unterschiedliche Wesen handele. Auch der Vergleich mit dem Volvo und Corvette hinke, weil beispielsweise ein Corvette immer die typischen Reifen und Bremsen der Corvette-Hersteller hat (und nicht die des Volvos). Männer und Frauen hingegen kann man zwar anhand ihrer Geschlechtsorgane differenzieren, nicht jedoch anhand ihrer kognitiven Fähigkeiten. Aus der räumlichen Wahrnehmung oder den Fähigkeiten in Mathematik alleine lässt sich aber nicht erkennen, ob wir es mit Männern und Frauen zu tun haben (vgl. FINE et al 2014).
Spiegelneuronen
Nichtsdestotrotz halten eine Reihe populärwissenschaftlicher Bücher am Mythos fest und suchen hierbei Verbindungen zu den Unterschieden bei Geschlechtern, die im Gehirn liegen. Hierzu zählen auch die sogenannten Spiegelneuronen. Hierbei handelt es sich um Nervenzellen die in unserem Gehirn beim Betrachten eines Vorgangs ein gleiches Aktivitätsmuster zeigen, wie bei dessen eigener Ausführung. Seit ihrer Beschreibung 1992 diskutiert man, ob Spiegelneuronen nicht nur bei der Nachahmung, sondern auch eine entscheidende Rolle bei der Empathie spielen (DE PILLIGRINO et al. 1992). Nachfolgende Forschungen haben jedoch keine Anhaltspunkte zwischen Spiegelneuronen und Empathie nachweisen können, vor allem weil Empathie nicht mit Nachahmung verwechselt werden darf (im Sinne einer Gefühlsansteckung) (vgl. z. B. HICKOK 2009, 2015, PASCOLO et al. 2010, HEYES 2010, KOSOGONOV 2012). Weiteres zu Spiegelneuronen findet sich hier

Abb. 6: Spiegelneuronen?
Da Frauen angeblich empathischer sind als Männer, sind natürlich die Spiegelneuronen aktiver als bei Männern, so das Vorurteil. So auch die Aussagen von Louann BRIZENDINE (2006), die ein populärwissenschaftlichen Buch herausbrachte (The Female Brain), welches hunderte wissenschaftlicher Artikel zitiert. Jedoch versagt dieses Buch darin wissenschaftliche Standards einzuhalten. YOUNG & BALABAN (2006) stellen in einem Review fest, dass das Buch durchsetzt ist mit wissenschaftlichen Fehlern bezüglich Gehirnentwicklung, dem Neuroendokrinen System und der Natur der Geschlechterunterschiede insgesamt. Außerdem werden die erwähnten „Fakten“ bei BRIZENDINE (2006) nicht in den zitierten Quellen unterstützt. FINE (2010) untersuchte die Behauptungen auf Seite 162 – 164 in BRIZENDINE (2006) Werk. So wird eine Studie von Psychotherapeuten zitiert (RAINGRUBER 2001) bei der Therapeuten in der Lage waren die Handlungen ihrer Patienten zu spiegeln. BRIZENDINE (2006, S. 162) erwähnt, dass die Therapeuten, die die Handlung ihrer Patienten spiegeln konnten allesamt Frauen waren und somit Frauen sich besser in die Lage anderer versetzen können. Leider vergisst sie nur zu erwähnen, dass bei dieser Studie nur Frauen teilnahmen, also ein Vergleich zwischen Geschlechtern gar nicht möglich war. BRIZENDINE (2006) nächste Behauptung ist, dass sich Mädchen besser in die Lage anderer versetzen können und zieht als Bewies Meta-Analysen von MCCLURE (2000) und HALL (1978, 1984) heran. Diese Studien ergaben zwar tatsächlich, dass Mädchen besser darin sind, sich in andere hineinzuversetzen, dieser Unterschied ist jedoch gering. MCCLURES (2000) Meta-Studie zeigt, dass 54% der Mädchen besser die Gesichtsausdrücke anderer nachvollziehen können als der Durchschnitt, bei Jungen liegt die Zahl bei 46% über dem Durchschnitt. HALLS Studien hingegen zeigen, dass wenn man beim „PONS nonverbal decoding task“ zufällig einen Jungen oder ein Mädchen aussucht, in einem Drittel der Fälle die Jungen besser abschneiden.
BRIZENDINE (2006, S. 162) übertreibt jedoch diese Unterschiede und sucht die Erklärung in der Funktion der Spiegelneuronen. Hierzu zitiert sie eine Studie (OBERMAN et al. 2005), bei der postuliert wird, dass das System der Spiegelneuronen beim Erahnen des Geisteszustand anderer Personen eine Rolle spielt. Doch in dieser Studie wurden nur Männer untersucht, einige von denen Autisten. Auf Seite 163 liest man bei BRIZENDINE (2006), dass bei Studien, die Aktivitäten des Gehirns bildlich darstellen (z. B. über fMRT), dass Frauen besonders gut beim spiegeln von Emotionen sind. Zur Unterstützung zitiert sie die Arbeit von SINGER et al. (2004), die die Hirnaktivität bei Probanden gemessen und verglichen haben, wenn eine Person einen schmerzhaften Elektroschock auf die Hand bekam oder eine nahestehende Person dabei zusehen musste. Man fand heraus, dass einige gleiche Hirnregionen, sowohl wenn man den Elektroschock bekam als auch dabei zusehen musste, aktiviert wurden. Das Problem der Studie: es wurde nur bei Frauen gemessen. Auch hier gab es also keinen Geschlechtervergleich, wodurch nicht daraus geschlossen werden kann, dass Frauen darin besser sind. Eine weitere Studie, die BRIZENDINE (2006) zur Unterstützung ihrer Behauptung, dass Frauen emotionaler sind, zitiert, ist jene von IDIAKA et al. (2002). Dort erhielten die Probanden Bilder von Gesichtern bei emotionalen Ausdrücken (positiv, negativ, neutral) und man sollte dabei das Geschlecht der Gesichter zuordnen. Die Studie verglich aber keine Geschlechter, sondern junge und alte Menschen miteinander. Unterschiede bei Geschlechtern wurden nicht erwähnt. BRIZENDINE (2006, S. 158) behauptet in Folge, dass Spiegelneuronen bei Frauen häufiger vorkommen, als bei Männern und zitiert hierzu mehrere Studien, die das bewiesen haben sollen: ORZHEKHOVSKIA (2005), UDDIN et al. (2005), OBERMAN et al. (2005) und OHNISHI et al. (2004). Die erste Studie befasste sich mit postmortalen Eigenschaften der Neuronen im Frontallappen. Die drei weiteren behandelten zwar einige Aspekte der Spiegelneuronen, aber keine von vergleich dabei Unterschiede zwischen den Geschlechtern, noch wurde über solche Unterschiede spekuliert.
Hippocampus und Amygdala
Der Hippocampus und Amygdala sind Teil des limbischen Systems, welches für die Verarbeitung von Emotionen eine Rolle spielt. Es ist spielt aber auch bei intellektuellen Leistungen eine Rolle (Abb. 7). Es wurde folgender Befund gemacht: Frauen haben einen größeren Hippocampus, Männer eine größere Amygdala (GOLDSTEIN et al. 2001). Der Hippocampus spielt u. a. eine Rolle bei Gedächtnisleistungen.
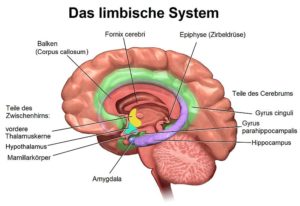
Abb. 7: Limbisches System
TAN et al. (2016) untersuchten 76 Publikationen, welche die Größe des Hippocampus von über 6000 Individuen gemessen haben. Ihr Ergebnis: die Unterschiede zwischen den Geschlechtern verschwinden.
Hier zeigt sich ebenfalls wieder der Unterschied zwischen Einzel-Studien und Meta-Analysen!
Wenn auch die Größe und das Volumen des Hippocampus zwischen den Geschlechtern zu vernachlässigen sind, so gibt es aber dennoch Unterschiede, die nicht zu vernachlässigen sind. Beispielsweise finden sich im Hippocampus eine hohe Konzentration an Rezeptoren für Geschlechtshormone (LENROOT & GIEDD 2010, WOOLEY 2007). Weiterhin wachsen die Zellen im Hippocampus von Frauen schneller als bei Männern (LENROOT & GIEDD 2010). CAHILL (2006) beschreibt zwar die Größenunterschiede des Hippocampus bei Männern und Frauen, die durch obige Meta-Analyse in Frage gestellt werden können. Er listet jedoch weitere Unterschiede auf: So ist das Volumen der CA1 Region des Hippocampus, inklusive der Anzahl der Pyramiden-Zellen (besonders große Nervenzellen) bei männlichen Ratten größer als bei weiblichen, wie auch die Dichte der Neuronen im Gyrus dentatus (im vorderen Bereich des Hippocampus). Auch das Neurotransmitter-System unterscheidet sich (MADEIRA & LIEBERMAN 1995). Die Rezeptor-Affinität für Glucocorticoide (Hormone aus der Nebennierenrinde) ist bei weiblichen Ratten nur halb so groß wie bei männlichen (TURNER & WEAVER 1985). Östrogen kann die Erregbarkeit der Zellen im Hippocampus verändern (TEYLER et al 1980), die Struktur bei Dendriten beeinflussen (COOKE & WOOLLEY 2005) und den Erinnerungsprozess modulieren (PACKARD et al. 1996). Der Hippocampus spielt auch eine Rolle bei Rückkopplungsprozessen von Östrogen eine Rolle und wird auch von Testosteron beeinflusst (JURASKA 1991). KOROL et al. (2004) entdeckten, dass Änderungen im Gleichgewicht von Geschlechtshormonen eine Verlagerung der Lernstrategien vom Hippocampus in Richtung Striatum begünstigen. Das Corpus striatum – kurz auch Striatum und auf Deutsch „Streifenkörper“ – ist der oberste Teil der Basalganglien und gehört zu den hochkomplexen motorischen Regelkreisen des Großhirns. JURASKA et al. (1985) konnten über entgegengesetzte Auswirkungen früher Erfahrungen in der Dendriten-Struktur im Gyrus dentatus ausfindig machen: weibliche Ratten, die in einer bereicherten Umgebung aufgewachsen sind, zeigten eine höhere Verzweigung bei den Dendriten als männliche. Eine weniger reiche Umgebung sorgte dafür, dass die Verzweigung der Dendriten bei Weibchen stärker degenerierte als bei Männchen. Diese Befunde erinnern an jene von SHORS et al. (2001), der ebenfalls den Hippocampus von Ratten untersuchte. Es zeigt erneut, wie stark Umweltfaktoren auf Nervenzellen einwirken können.
Es gibt weitere Hinweise darauf, dass es im Bereich von Neurotransmittern Unterschiede zwischen Geschlechtern gibt; so bei der Monoamino-Oxidase bei Menschen und Ratten (CURTIS et al. 2005, ROBINSON et al. 1977) und bei der Serotonin-Synthese (NISHIZAWA et al. 1997), nebst anderen (CAHILL 2006). Auch der Menstruationszyklus hat Auswirkungen auf die Hormonlevel und Botenstoffe im Gehirn (CAHILL 2006, Box 1).
CAHILL (2006) zitiert hier im wesentlichen Studien, die an Ratten durchgeführt wurden. Zweifelsohne lassen sich auch hier einige Beobachtungen bei menschlichen Gehirnen machen. Alleine die Tatsache, dass sich im Hippocampus unterschiedliche Rezeptoren für Hormone befinden, deutet darauf hin, dass es biochemische Unterschiede geben muss. Für die Medizin und die Verwendung von Präparaten, die häufig mit dem Hormon-Rezeptor-System in Einklang gebracht werden müssen, spielen diese Befunde sicherlich eine Rolle. Hier ist aber auch deutlich anzumerken, dass unser Gehirn nicht nur auf die Umwelt reagiert, sondern von ihr auch modifiziert wird. Es ergibt sich eine dialektische Wechselwirkung zwischen sozialem, Umwelt, Gehirn, Hormonen und Genetik. Es lassen sich hieraus aber auch keine intellektuellen Fähigkeiten direkt ableiten, zum einen weil z. B. weniger Nervenzellen durch die Produktion von mehr Rezeptoren oder Neurotransmittern kompensiert werden können, zum anderen sind die einzelnen Hirnregionen niemals isoliert zu betrachten, sondern bedingen ebenso einander, sodass eine Hirnregion niemals alleine für alle kognitiven Prozesse zuständig ist. Beispiele in den anderen Abschnitten haben dies hoffentlich verdeutlichen können. Dass sich aus biochemischen oder morphologischen Unterschieden im Hippocampus oder anderen Gehirnregionen keine kognitiven Fähigkeiten ableiten lassen, gibt sogar Larry CAHILL (2005) zu, der feststellt, dass bis heute keine Beweise dafür gefunden wurden, dass anatomische Unterschiede dazu führen, dass Frauen keine hohen Positionen in Mathematik, Physik und Ingenieurswissenschaften haben. Denn eine Korrelation zwischen anatomischen Unterschieden im Gehirn und intellektuellen Fähigkeiten bestünde nicht. Bei Intelligenztests schneiden Frauen im Durchschnitt genauso ab wie Männer, trotz ihrer Unterschiede, was dazu führt, dass beide dieselben Aufgaben auf andere Weise durchführen können (CAHILL 2005, S. 40 & S. 47 Infobox). Aber – und das zeigt die Erfahrung der bisherigen Befunde – man sollte niemals die Überlappungen zwischen den Geschlechtern vergessen, die wesentlich größer sind, als die Unterschiede.
Wie stark auch der Hippocampus durch Umwelteinflüsse verändert werden kann, zeigte eine Studie, die die Größe des Hippocampus bei 9 und 10 jährigen Jungen und Mädchen analysierten. Sie entdeckten, dass es einen Zusammenhang zwischen physischer Fitness und Hippocampus-Größe gibt: die fitteren hatten einen größeren Hippocampus, unabhängig davon, ob sie Junge oder Mädchen waren. Sie waren auch besser beim Gedächtnistraining (CHADDOCK et al. 2010). Sich geistig und körperlich fit zu halten hat tatsächlich mehr Einfluss auf das Gehirn als das angeborene Geschlecht. Beispielsweise fand eine Studie heraus, dass Londoner Taxifahrer einen größeren Hippocampus (genauer gesagt die Graue Substanz im Hippocampus) haben, da sie sich die 25.000+ Straßen der Stadt merken müssen (und das bei all dem Verkehrschaos) (WOLLET & MAGUIRE 2011). Eine weitere Studie (KELLER & JUST 2016) belegten, dass kurzes räumliches Navigationstraining das Gehirngewebe einer Person verändert und die Kommunikation des veränderten Gehirngewebes mit anderen Gehirnbereichen verbessert, die an der räumlichen Navigation beteiligt sind. Diese bahnbrechende Entdeckung stellt eine kritische Verbindung zwischen struktureller Größe und funktionellen Konnektivitätsveränderungen des Gehirns her, die im Hippocampus beim räumlichen Lernen auftreten. Diese Studie zeigt auch, dass die mit räumlichem Lernen verbundenen Gehirnveränderungen davon abhängen, wie neuronale Aktivität die Kommunikation zwischen dem Hippocampus und anderen für die Navigation erforderlichen Regionen synchronisiert. Tatsächlich hat auch Armut einen Einfluss auf die Gehirnaktivität. Mehrere Studien belegen, dass Armut, besonders bei Kindern, nachteilige Auswirkungen auf die Gehirnstruktur hat (z. B. NOBLE et al. 2015, MACKEY et al. 2015, HAIR et al. 2015). Familiäre Probleme (von haben ebenfalls einen Einfluss auf die Gehirnentwicklung (vgl. z. B. WALSH et al. 2014 zu den negativen Auswirkungen auf das Kleinhirn und die Graue Substanz). Kognitives Training kann dabei die Gehirnleistung hingegen verbessern – auch bei in Armut aufgewachsenen Kindern (GAMIO et al. 2014). Ebenfalls hat Stress negative Auswirkungen auf den Hippocampus (CHETTY et al. 2014)
Wenn also einer die waghalsige Behauptung aufstellt, dass arme Menschen „dumm“ sind, so ist dies zweifelsohne eine Konsequenz der sozioökonomischen Verhältnisse und nicht der Menschen an sich. Dummheit ist somit kein Schicksal – und Armut genauso wenig, wenn dagegen gekämpft wird! Es zeigt sich also, dass gesellschaftliche Phänomene mehr Einfluss auf das Gehirn haben, als das Geschlecht. Das hat zumindest zur Konsequenz, dass der Kampf gegen Armut (und Klassenunterschiede) einen deutlich wichtigeren Stellenwert einnimmt, als eine „gendergerechte“ Sprache.
Die Amygdala spielt u. a. eine Rolle bei Emotionen wie Angst und Wut. Nun haben aber Meta-Studien zu beiden Strukturen den Größenunterschied beider Strukturen relativiert.
Eine Meta-Analyse von MARWHA, HALARI & ELIOT (2017) ergab, dass die Unterschiede nicht signifikant sind. Man untersuchte 46 Studien, die das Volumen der Amygdala von 6726 Individuen beiden Geschlechtes. Man fand heraus, dass das Volumen der Amygdala bei Männern etwa 10 % größer ist, was etwa dem 10% größeren Volumen des Gesamtgehirns entspricht.
SAX (2006) zitiert mehrere Studien in Bezug zur Amygdala, die zeigen sollen, dass negative Emotionen bei Männern hauptsächlich von der Amygdala gesteuert werden, bei Frauen hingegen in der Hirnrinde. Eine dieser Studien (SCHNEIDER et al. 2000) untersuchte 13 Männer und 13 Frauen bei induzierter Trauer und stellen tatsächlich fest, die die rechte Amygdala bei Männern stärker aktiviert war (bei Frauen hingegen war die linke Amygdala aktiver; bei Freude zeigten sich keine Geschlechtsunterschiede). Die Autoren diskutieren in ihrer Arbeit jedoch keine Geschlechterunterschiede. Eine weitere Studie (KILLGORE & YURGELON-TODD 2001) untersuchte die Aktivität der Amygdala beim Betrachten ängstlicher und erfreuter Gesichter und fand neben der Tatsache, dass bei Männern die rechte Amygdala aktiver war und bei Frauen dafür die linke keine Geschlechtsunterschiede. Es gab lediglich einen Lateralisations-Effekt, aber keinen Unterschied an der Amygdala-Funktion an sich (die Hirnrinde wurde nicht untersucht). SAX (2006) zitiert auch die Metastudie von WAGNER et al. (2003) als Beweis dafür, dass Emotionen bei Männern und Frauen unterschiedlich aktiviert werden. Jedoch stimmen ihre Ergebnisse mit der Meinung von SAX nicht überein, dass Emotionen bei Frauen hauptsächlich in der Hirnrinde und bei Männern unterhalb der Hirnrinde (subcortical) verarbeitet werden. Die Studie von WAGNER et al. (2003) berichtet, dass bei Männern hierbei hauptsächlich der hintere sensorische und assoziative Cortex, der untere frontale Cortex (alle drei Bestandteile der Hirnrinde) und der dorsale Striatum (unterhalb der Hirnrinde) häufiger aktiviert werden als bei Frauen. Bei Frauen hingegen wird der mediale frontale Cortex, der Thalamus und das Kleinhirn stärker aktiviert (Thalamus und Kleinhirn gehören nicht zur Hirnrinde). Auch hier ist nicht auszuschließen, dass unterschiedliche Hirnregionen einander kompensieren können und ähnliche Aufgaben erfüllen. Übrigens wird die Amygdala nicht nur bei negativen Emotionen aktiviert, sondern z. B. auch Bilder von lachenden Menschen betrachtet (CUNNINGHAM & KIRKLAND 2013).
Hypothalamus
Beim Hypothalamus handelt es sich um eine wichtige Struktur im Gehirn, welches bei der Regulation vieler biologischer und psychologischer Funktionen beteiligt ist. Er ist mit der Hypophyse verbunden, welches die zentrale Schaltstelle der Hormonproduktion ist. Erhält die Hypophyse vom Hypothalamus entsprechende Signale, werden von der Hypophyse Hormone ausgeschüttet, die die entsprechenden Hormondrüsen (z. B. Hoden, Ovarien, Nebennierenrinde, Bauchspeicheldrüse, Schilddrüse) erreichen, die wiederrum ihre spezifischen Hormone (z. B. Testosteron, Östrogen, Corticoide, Insulin, Thyroxin) ins Blut abgeben. Das Hormonsystem verfügt über einen Rückkopplungsmechanismus, dass wenn z. B. genügend Testosteron produziert wurde, chemische Botenstoffe zurück zum Hypothalamus befördert werden, des dadurch seine Stimulation der Hypophyse hemmt (vgl. Abb. 8). Der Hypothalamus steht natürlich unter Kontrolle des Gehirns und seine Aktivität steht im Zusammenhang mit genetischen Faktoren wie Umweltfaktoren.
Abb. 8: Rückkopplungsmechanismus des Hormonsystems. Quelle
Das Hormonsystem als solches ist relativ komplex und beeinflusst die Entwicklung auf unterschiedliche Weise. Wir werden uns damit gesondert nochmal näher befassen. Da der Hypothalamus so stark in das Hormonsystem integriert ist, gibt es natürlich auch hier entsprechende Unterschiede: Da er z. B. auch den Menstruationszyklus der Frau steuert, wie auch die Ausbildung der Spermien liegt hier schon mal biochemisch ein Sexualdimorphismus vor. Der Hypothalamus hat auch bei Emotionen eine wichtige Aufgabe. Eine bestimmte Zellgruppe des Hypothalamus (INAH-3) ist bei Männern größer. Interessanterweise ist diese Zellgruppe durchschnittlich bei homosexuellen Männern kleiner als bei heterosexuellen (LEVAY 1991), wobei eine Studie von BYNE et al. (2001) festgestellt hat, dass trotz des Volumenunterschiedes zwischen hetero- und homosexuellen Männern sich kein Unterschied in der Anzahl der Neuronen findet. Man weiß aber nicht so recht, wie stark die INAH-3-Region des Hypothalamus mit der sexuellen Orientierung korreliert. Man geht eher davon aus, dass diese eher eine Rolle beim Menstruationszyklus spiele (JORDAN-YOUNG 2010, S. 50). SAVIC, BERLUND & LINDSTRÖM (2005) entdeckten, dass durch den Geruch von Pheromonen aus Männerschweiß der Hypothalamus bei heterosexuellen Frauen und homosexuellen Männern stärker reagierte. Der Hypothalamus hetero- und homosexueller Männer reagiert auch unterschiedlich auf das Antidepressivum Fluoxetin (KINNUNEN et al. 2004).
Die BSTc-Region des Hypothalamus spielt eine Rolle beim männlichen Sexualverhalten und ist bei ihnen daher etwa doppelt so groß wie bei Frauen und hat etwa doppelt so viele Neuronen. Einige Untersuchungen haben gezeigt, dass bei jenen Männern, die sich zur Frau operieren haben lassen, die BSTc-Region denen von Frauen ähnlich ist, wobei hormonelle Einflüsse angeblich keine Rolle spielten (KRUJIVER et al. 2000, ZHU et al. 1995). KRUJIVER et al. (2000) sowie ZHU et al. (1995) untersuchten dabei das Gehirn von sechs transsexuellen Menschen.
Es hat damit den Anschein, dass der Hypothalamus eine Rolle bei der sexuellen Orientierung spielt. Das sollte nachdem, was man über den Hypothalamus, der auch in der Pubertät stärker aktiv wird und z. B. auch den Menstruationszyklus steuert, nicht verwundern. Es wäre aber hier dennoch Vorsicht geboten, die sexuelle Orientierung alleine am Hypothalamus festzulegen. Zum einen wissen wir, dass unser Gehirn, und damit auch der Hypothalamus, auf Umwelteinflüsse reagiert, zum anderen ist menschliches Sexualverhalten zu komplex, als dass es auf nur einen Faktor reduziert werden kann. Das trifft auch für die BSTc-Region und Transsexualität zu. Denn mittlerweile gibt es eine Reihe an Studien, die die Theorien des Transsexuellen-Gehirns“ (und auch des „Schwulen-Gehirns“) ablehnen. Es soll zwar nicht abgestritten werden, dass diese Region des Hypothalamus eine Rolle im Sexualverhalten hat: doch auch hier sind die Überschneidungen zwischen den Geschlechtern vorhanden, es lässt sich also nicht behaupten, dass die BSTc-Region bei jedem Mann größer ist als bei Frauen. Außerdem handelt es sich bei KRUJIVER et al. (2000) sowie ZHU et al. (1995) um Postmortal-Studien, also nach dem Tode der Patienten. Beide Arbeiten untersuchten das Gehirn von sechs bis acht transsexuellen Personen, die Stichprobe war also sehr gering. Wir wissen mittlerweile, dass auch Alter und Tod einen Einfluss auf die Hirngröße und ihren Abbau haben.
CHUNG, DE VRIES & SWAAB (2002) konnten zeigen, dass der sexuelle Dimorphismus der BSTc-Region erst im Erwachsenenalter auftritt, wodurch die Theorie, dass Transsexuelle schon in ihrer Kindheit einen anderen Hypothalamus haben, ins Wanken gerät. Transsexuelle berichten, dass sie schon während der Kindheit sich im falschen Geschlecht fühlten (z. B. LAWRENCE 2003). CHUNG, DE VRIES & SWAAB (2002) gehen entsprechend davon aus, dass Hormone einen Einfluss auf die Geschlechtsidentität haben können, wodurch sich die BSTc-Region nachträglich ändere. Zwar schließen KRUJIVER et al. (2000) und ZHU et al. (1995) hormonelle Einflüsse der untersuchten Gehirne in Bezug zur BSFc-Region aus, doch LAWRENCE (2007) geht stark davon aus, dass die Änderung dieser Region Folge der Hormontherapie war. So konnten HULSHOFF POL et al. (2006) belegen, dass die Behandlung von Transgender-Personen mit Hormonen einen Einfluss auf das Gehirnvolumen der Patienten hat. Dabei nahm auch das Volumen des Hypothalamus ab, wodurch die Mann-zu-Frau-Transgender-Personen entsprechend auch eine kleinere BSTc-Region haben. Das entspricht auch den Befunden der Postmortal-Studien. KRUJIVER et al. (2000) irren, wenn sie die Behauptung aufstellen, dass Hormone die BSTc-Region nicht beeinflussen. LAWRENCE (2006) schlussfolgert, dass die Theorie von einem Transsexuellen-Hirn den klinischen Befunden nicht entspreche und man neue Wege finden müsse, dieses Phänomen zu erklären.
Korrelieren Gehirnstruktur und Psyche miteinander?
Dass es wahrscheinlich keine „besondere“ Transsexuellen- oder Schwulen-Struktur bei menschlichen Gehirnen gibt, heißt natürlich nicht, dass sexuelle Identität und Orientierung „frei wählbar“ sind und man durch „richtige Erziehung“ alle schön zu Heterosexuellen machen kann. Unser Gehirn arbeitet auf eine völlig andere Weise: Es ist plastisch, anpassungsfähig und der permanente Abbau und Aufbau von Nervenverknüpfungen lassen eine direkte Übertragung von Gehirnstruktur zum Verhalten nicht zu. Seine Funktionsweise wurde im Kapitel „Gehirn und Umwelt“ ausreichend beschrieben. Persönlichkeitsmerkmale wie z. B. Intelligenz lassen sich auch nicht an Gehirnstrukturen messen. Zwar gibt es einige Studien, die gewisse Persönlichkeitsmerkmale anhand von Gehirnstrukturen erkannt haben wollen: NOSTRO et al. (2017) wollen entdeckt haben, dass die Menge an grauer Hirnsubstanz im Sulcus parietooccipitalis, einer Großhirnfurche zwischen Scheitel- und Hinterhauptslappen, bei männlichen Teilnehmern tatsächlich Hinweise darauf zu liefern scheint, wie extravertiert, gewissenhaft und emotional stabil diese sind. Interessanterweise fanden sie aber keine Korrelation bei Frauen, was sie – welch Wunder! – auf weibliche Hormone zurückführen (ohne das zu belegen). DEYOUNG et al. (2010) meinen einen Zusammenhang zwischen Gehirnstruktur und den „Big Five“ der Psychologie (Extraversion, Neurotizismus, Verträglichkeit, Gewissenhaftigkeit, Offenheit für Erfahrungen) gefunden zu haben. Ihnen zur Folge korreliert Extraversion mit dem Volumen des orbifrontalen Cortex, Neurotizismus mit dem Gehirnregionen, die man mit negativen Erlebnissen assoziiert, Verträglichkeit mit dem Volumen in den Regionen, die Informationen über die Absichten und den Geisteszustand anderer Personen verarbeiten und Gewissenhaftigkeit korreliere mit dem Volumen im lateralen präfrontalen Kortex, einer Region, die an der Planung und der freiwilligen Kontrolle des Verhaltens beteiligt ist.
Es gibt jedoch bei solchen Vorstellungen eine Reihe von Problemen:
- Zuallererst ist anzumerken, dass Korrelation zwischen zwei Merkmalen nicht automatisch Ursache und Wirkung bedeutet. Sollte aber ein Zusammenhang bestehen, ist auch nicht die Frage geklärt, ob gewisse Gehirnstrukturen diese Merkmale produzieren, oder ob das Verhalten (bzw. die psychologischen Merkmale) diese Gehirnstruktur formen. Wir wissen ja, dass beides geht.
- Die Autoren haben den phrenologischen Rahmen buchstäblich wieder übernommen, wobei eine größere lokale Gehirnregion mehr lokale Nervenstärke und mehr Ressourcen für das bedeutet, was in der Region kontrolliert wird. Das stammte aus einer Zeit, als das Gehirn mit einem Muskel verglichen wurde und man daher dachte: Mehr Masse = Mehr Hirn = Mehr Nerventätigkeit. Daher wurden ja die größeren Männergehirne als überlegen angesehen. Heute wissen wir, wie hier auch schon mehrmals dargestellt, dass größer nicht automatisch mehr und schon gar nicht besser bedeutet.
- Auch wenn es wahr ist, dass ein größerer Bereich des Gehirns mehr Neuronen erhalten kann, variiert die Dichte der Grauen Substanz im gesamten Gehirn und ändern sich zudem im Verlauf des Lebens in unterschiedlichen Geschwindigkeiten, sodass Regionen mit unterschiedlichen Größen tatsächlich zu einem bestimmten Zeitpunkt die gleiche Anzahl von Neuronen und zu einem anderen Zeitpunkt eine unterschiedliche Anzahl enthalten können.
- Es scheint völlig vernünftig zu sein anzunehmen, dass mehr Neuronen mehr Rechenleistung implizieren. Die Beziehung von Funktion zur Struktur ist jedoch subtiler. Obwohl die Funktion durch Neurogenese verfeinert werden kann, indem mehr Neuronen hinzugefügt werden, kann sie sowohl durch Hinzufügen als auch durch Beschneiden von neuronalen Verbindungen verfeinert werden. Abhängig von der auszuführenden Funktion kann das Erhöhen der Population oder der Dichte von Neuronen vorteilhaft sein oder schädlich sein. Hier gibt es wirklich keine gute Regel.
- Auch wenn eine Region aufgrund ihrer Größe bei einer bestimmten Person mehr Neuronen enthält, bedeutet dies nicht, dass diese Person mehr Neuronen verwendet, um die Funktionen auszuführen, die dieser Region in Studien zugeordnet wurden. Eine Person mit mehr neuronalen Ressourcen in einem bestimmten Bereich kann den „Überschuss“ für verschiedene Zwecke verwenden. Ohne gute Funktionsstudien dieser Personen ist es unmöglich das zu wissen.
- Auch wenn ein Individuum mehr Neuronen für eine bestimmte Aufgabe verwendet, bedeutet dies nicht, dass diese Neuronen im Gehirn des Individuums „einflussreicher“ sind (und somit einen größeren Einfluss auf das Verhalten haben). Wie „einflussreich“ ein bestimmter neuronaler Bereich ist, hängt weit mehr von den Details ihrer Synchronität und ihrer Konnektivität ab. Außerdem kann ein Individuum mehr Ressourcen für ein Problem aufwenden, wenn es relativ schwieriger ist, was eher auf relative Schwäche als auf relative Stärke hinweist. Es gibt suggestive Beweise in der Literatur, die die Komplexität dieser Gleichung belegen. Londoner Taxifahrer scheinen in einem bestimmten Teil ihres Hippocampus ein größeres Volumen an grauer Substanz zu haben (verglichen mit Londoner Busfahrern) (WOLLET & MAGUIRE 2011)). Auf der anderen Seite gibt es eine Fülle von Literatur, die darauf hindeutet, dass Experten oft weniger neuronale Ressourcen für Probleme einsetzen als Neulinge.
- einen Teil der Studien scheinen die Autoren falsch verstanden zu haben. DEYOUNG et al. (2010) zitieren das Beispiel, dass die Dichte der Grauen Substanz bei älteren Menschen nach dem lernen des Jonglierens zunimmt, es gibt jedoch keine Volumenänderungen (BOYKE et al. 2008). Sie zitieren eine weitere Studie (MCDANIEL 2005), bei der eine positive Korrelation zwischen Gehirnvolumen und IQ festgestellt wurde. Nichts in dieser Studie spricht jedoch für die Annahme, dass lokale anatomische Unterschiede mit spezifischen Verhaltens- und Funktionsunterschieden korrelieren.
- Einige ihrer funktionellen Annahmen sind ebenfalls fehlerhaft: Sie ordnen bestimmten Hirnarealen Persönlichkeitsmerkmale zu. Es sind aber viele Hirnregionen an jedem kognitiven Prozess beteiligt. Selbst so einfache Dinge wie das Schreiben am Computer erfordert, dass die Hirnareale miteinander kooperieren: jene Neuronen, die ihre Finger- und Handbewegungen steuern, das Sprachzentrum, ihr Erinnerungsvermögen usw. usf. sind dabei beteiligt. Warum sollte es da bei psychologischen Merkmalen anders aussehen? Die höhere Nerventätigkeit erfordert, dass verschiedene Hirnareale miteinander kooperieren und die Summe dieser Nerventätigkeit produziert als Qualitätssprung unser Verhalten. Unser entwicklungsgeschichtlicher Aspekt (unsere individuelle Erfahrungen sowie unsere Stammesgeschichte) tun ihr Übriges. Es ist daher prinzipiell gar nicht möglich, dass gewisse Persönlichkeitsmerkmale an einer Gehirnstruktur gefunden werden können, da eben unser Gehirn nicht so funktioniert!
- Natürlich sind entsprechende Hirnareale für eine „normale“ psychologische Tätigkeit notwendig. Werden diese Areale irreparabel beschädigt kommt es zur Störung der Psyche. Das ist jedoch eine triviale Feststellung: Ohne Gehirn können wir nicht denken. Doch längst nicht jede psychologische Störung – und damit schon gar nicht die psychologischen und kognitiven Eigenschaften „gesunder Menschen“ – sind auf Störungen bzw. Änderungen der Hirnanatomie zu reduzieren. Der sowjetische Neurobiologe und Psychologe Iwan Pawlow unterschied beispielsweise bei Geisteskrankheiten zwischen organischen Störungen (Areale der Gehirnstruktur sind irreparabel geschädigt) und funktionellen Störungen (funktionelle Tätigkeit des neuronalen Netzwerks). Letztere hielt Pawlow für heilbar (vgl. WELLS 1989, S. 260). Ich fühle mich nicht kompetent genug, um zu urteilen, ob alle funktionellen Geistesstörungen heilbar sind. Aber Pawlows Darstellung hat einen Vorteil: Er liefert einen Hinweis darauf, dass unsere Persönlichkeit nicht an einer Gehirnstruktur ablesbar ist, da diese von der Tätigkeit des gesamten Nervensystems abhängen.
Tatsächlich ist die Vorstellung Persönlichkeiten anhand von Gehirnmerkmalen festzustellen nicht nur wissenschaftlich falsch, sondern politisch reaktionär. Dies hat der Erste Teil unseres Gehirn-Kapitels wunderbar belegt.
Dass es Geschlechterunterschiede im Gehirn, bzw. in der Gehirnfunktion gibt soll nicht abgestritten werden, beispielsweise regelt das Gehirn unser Hormonsystem und die Keimzellenproduktion – und bisher hat noch kein Mann menstruiert oder wurde schwanger und Frauen ejakulieren nicht. Aber die Differenzen sind gering, es gibt große Überschneidungen und Persönlichkeitsmerkmale lassen sich nicht auf gewisse Hirnstrukturen reduzieren, weiterhin reagiert unser Gehirn aktiv auf die Umwelt und verändert sich, so dass man nicht pauschalisierend sagen kann, dass unser die Unterschiede angeboren und unveränderbar sind. Aber selbst wenn dies der Fall wäre: Das Anerkennen von Unterschieden ist an sich kein Sexismus. Aus der Tatsache dass Männer durchschnittlich ein größeres Gehirnvolumen haben, kann man nicht schlussfolgern, dass Männer „schlauer“, „stärker“, „intelligenter“ oder „besser“ wären. Selbiges würde für die Beobachtung gelten, dass Frauen „emotionaler“ seien, daraus haben Männer noch längst nicht ein Herz aus Stein. Ein kennenlernen, wo es tatsächlich Unterschiede gibt (und welche Ursachen sie haben könnten) kann auch insofern sinnvoll sein, dass man gegen Stereotypie, Vorurteile und Unterdrückung argumentativ vorgehen kann. Im dritten Teil der Gehirn-Saga der Geschlechter wollen wir uns daher mit einigen angeblichen und tatsächlichen kognitiven Unterschieden zwischen den Geschlechtern auseinandersetzen.
Literatur
ALLEN, DAMASIO & GRABOWSKI (2002): Normal neuroanatomical variation in the human brain: An MRI‐volumetric study. American Journal of Physical Anthropology Band 118, Nr. 4, 1., S. 341–358
BARON-COHEN (2004): The essential difference: Male and female brains and the truth about autism . New York : Basic Books
BEAN (1906): Some racial pecularities of the Negro Brain. American Journal of Anatomy 5: 353 – 432
BECK (2014): Hirnrissig. Die 20,5 größten Neuromythen – und wie unser Gehirn wirklich tickt. München: Hanser
BIRD et al. (2004): The impact of extensive medial frontal lobe damage on „The Theoriy of Mind“ and cognition. Brain, 127, 914 – 928
BISHOP & WAHLSTEIN (1997): Sex differences in the human corpus callosum: Myth or Reality? Neuroscience and Bihavoioural Reviews 21 (5): 581 – 601
BISWAL et al. (2010): Toward discovery science of human brain function. Proceedings of the National Academy of Sciences, 107(10), 4734–4739.
BLAKEMORE et al. (2007): Adolescence development of the neural circuitry for thinking about intentions. Social Cognitive and Affective Neuroscience, 2, 130-139
BLEIER (1986): Sex differences research: Science or belief? In BLEIER (1986, Hrsg.): Feminist approaches to science (S. 147-164). New York: Pergamon Press
BLUMBERG (2005): Basic Instinct – The genesis of behavoior. Thunder’s Mouth Press
BOLES (2005): A large-sample study of sex differences in functional cerebral lateralization . Journal of Clinical and Experimental Neuropsychology , 27 , 759 – 768 .
BOYKE et al. (2008): Training-induced brain structure changes in the elderly. Journal of Neuroscience. 2008; 28:7031–7035.
BRIZENDINE (2006): The Female Brain. London: Bantan Press
BROOKS-GUNN et al. (1992): Effects of early intervention on cognitive functions of low birth weight preterm infants . Journal of Pediatrics , 120 , 350 – 359
BUTTON et al. (2013): Power failure: why small sample size undermines the reliability of neuroscience. Nature Reviews Neuroscience volume 14, pages 365–376
BURACAS, FINE & BOYNTON (2005): The realtionship between task performance and functional magnetic resonance imaging response. Journal of Neuroscience 25 (12), 3023 – 3031
BURNETT et al. (2009): Development during adolescence of the neural processing of social emotion. Journal of Cognitive Neuroscience, 21 (9), 1736 – 1750
BYNE et al. (2001): The interstitial nuclei of the human anterior hypothalamus: an investigation of variation with sex, sexual orientation, and HIV status. Horm Behav. 2001 Sep;40(2):86-92.
CAHILL (2003): Sex- and Hemisphere-Related Influences on the Neurobiology of Emotionally Influenced Memory. Progress in Neuro-Psychopharmacology and Biological Psychiatry 27 (8): 1235–1241.
CAHILL (2005): His Brain, Her Brain. Scientific American
CAHILL (2006): Why sex matters for neuroscience . Nature Reviews Neuroscience , 7 , 477 – 484.
CAHILL (2014): Fundamental sex difference in the human brain architecture. PNAS January 14, 2014 111 (2) 577-578
CAHILL (2014a): Equal ≠ the same: Sex differences in the human brain. Cerebrum
CAROTHERS & REIS (2013): Men and women are from Earth: Examining the latent structure of gender. Journal of Personality and Social Psychology, 2013. 104(2): p. 385-407.
CHADDOCK et al. 2010): A neuroimaging investigation of the association between aerobic fitness, hippocampal volume, and memory performance in preadolescent children. Brain Res. 2010 Oct 28;1358:172-83.
CHETTY et al. (2014): Stress and glucocorticoids promote oligodendrogenesis in the adult hippocampus. Molecular Psychiatry volume 19, pages 1275–1283
CHUNG, DE VRIES & SWAAB (2002): Sexual differentiation of the bednucleus of the stria terminalis in humans may extend into adulthood. Journal ofNeuroscience, 22, 1027-1033.
COHEN (1988): Statistical Power Analysis for the Behavioral Sciences. 2. Auflage. Lawrence Erlbaum Associates, Hillsdale
COOKE & WOOLLEY (2005): Gonadal hormone modulation of dendrites in the mammalian CNS. J. Neurobiol. 64, 34–46
CORNEL (2014): Review: Ingalhalikar, M. et al. (2014) Article Review PSYC 747 11/26/2014
CUNNINGHAM & KIRKLAND (2013): The joyful, yet balanced amygdala: moderated responses to positive but not negative stimuli in trait happiness. Soc Cogn Affect Neursosci, doi 10.1093
CURTIS et al. (2005): Sexually dimorphic responses of the brain norepinephrine system to stress and corticotropin-releasing factor. Neuropsychopharmacology 31, 544–554
DEL GUIDICE et al. (2016): Joel’s et al. Method systemtically fails to detect large, consistent sex differences. Proceedings of the National Acadmiy of Sciences, 113 (14), E1965
DE PILLIGRINO et al. (1992): Understanding motor events: a neurophysiological study. Exp Brain Res. 1992;91(1):176-80.
DE VRIES & FORGER (2015): Sex differences in the brain: A whole body perspective. Biology of Sex Differences, 6 (1), 1-15
DE VRIES & SÖDERSTEN (2009): Sex differences in the brain: The relation between structure and function. Hormones and Behavior, 55 (5), 598 – 596
DEYOUNG et al. (2010): Testing Predictions From Personality Neuroscience: BrainStructure and the Big Five. Psychol Sci.; 21(6): 820–828.
DIETRICH & KANSO (2010): A review of EEG, ERP, and neuroimaging studies of creativity and ionsight. Psychol Bull, 136, (5): 822-848
ELIOT (2009): Pink brain blue brain. How small differences grow into troublesome gaps and what we can do about it . Boston, New York : Houghton Miffl in Harcourt
FINE (2010): Delusion of Gender. The real science behind sex differences. London: Icon Books
FINE (2014): Neuorsexism: A conversation with Cordelia Fine
FINE et al. (2013): Plasticity, plasticity, plasticity … and the rigid problem of sex. Trends in Cognitive Sciences Vol. 17 (11), 550-551
FINE et al. (2014): Reaction to “Equal ≠ The Same: Sex Differences in the Human Brain”, Cerebrum
FINK et al. (2009): Brain correlates underlying creative thinking: EEG alpha activity in professional vs. Novice dancers. Neuroimage, 46 (3): 854 – 862
FJELL, WALHOVD & FENNEMA-NOTESTINE (2009): Minute effects of sex on the aging brain: A multisample magnetic resonance imaging study of healthy aging and Alzheimer’s disease . Journal of Neuroscience , 29 , 8777 – 8783 .
FJELL et al. (2009): Minute effects of sex on the aging brain: A multisample magnetic resonance imaging study of healthy aging and Alzheimer’s disease . Journal of Neuroscience , 29 , 8777 – 8783
FOX, LEVITT & NELSON (2010): How the timing and quality of early experiences infl uence the development of brain architecture . Child Development , 81 , 28 – 40
FRASIER & YASNITSKY (2015): Deconstructiong Vygotsky’s victimization narrative: A re-examination of the ‚Stalinist supression‘ of Vygotskian theory. History of Human Sciences, 28 (2), 128-153
Freidenker Heft Nr. 3. 2018 „Menschenbild – Marxistische Persönlichkeitstheorie: Individuum und Gesellschaft“.
FRISTON & PRICE (2001): Dynamic representations and generative models of brain function. Brain Resarch Bulletin, 54 (3), 275 – 285
GAMIO et al. (2014): Enhancing inferential abilities in adolescence: new hope for students in poverty. Front. Hum. Neurosci., 09 December 2014
GESCHWIND & BEHAN (1982): Left-handedness: Association with immune disease, migraine and developmental lerning disorder. Proceedings of the National Academ of Sciences USA, 79, 5097-5100
GESCHWIND & GALABURDA (1985): Cerebral lateralization. Biological mechanisms, associations, and pathology: I. A hypothesis and a program for research JAMA Neurology 42(5):428-59
GETTLER et al. (2011): Longitudinal evidence that fatherhood decreases testosterone in human males. Proc. Natl. Acad. Sci. U.S.A. 108: 13194-13199
GIANAROS et al. (2007): Perigenual anterior cingulate morphology covaries with perceived social standing. Social Cognitive and Affective Neuroscience, 2(3), 161–173.
GIEDD et al. (2006): Puberty-related influences on brain development. Molecular and Cellular Endocrinology, 254/255, 154 – 162
GILLMORE et al. (2007): Regional Gray Matter Growth, Sexual Dimorphism, and Cerebral Asymmetry in the Neonatal Brain. Journal of Neuroscience 7 February 2007, 27 (6) 1255-1260;
GOLDSTEIN et al. (2001): Normal sexual dimorphism ot the adult human brain assessed by in vivo magnetic resonance imaging, Cereb Cortex, 11 (6): 490-497
GREENOUGH, BLACK & WALLACE (1987): What’s special about development? Thoughts on the bases of experience-sensitive synaptic plasticity. In W. T. Greenough & J. M. Juraska (Eds.), Developmental neuropsychobiology (pp. 387 – 407 ). Orlando : Academic Press
GUR et al. (1991): Gender differences in age effect on brain atrophy measured by magnetic resonance imaging . Proceedings of the National Academy of Sciences of the USA , 88 , 2845 – 2849
HACKMAN & FARAH (2009): Socioeconomic status and the developing brain. Trends in Cognitive Sciences, 13(2), 65–73.
HAIER et al. (1992): Regional glucose metabolic changes after learning a complex visuospatial/motor task: A positron emission tomographic study. Brain Research, 570, 134 – 143
HAIER et al. (2005): The neuroanatomy of general intelligence: sex matters. Neuroimage, 25, 320 – 327
HAIR et al. (2015): Association of Child Poverty, Brain Development, and Academic Achievement. JAMA Pediatr. 2015;169(9):822-829.
HALL (1978): Gender effects in deconding nonverbal cues. Psychological Bulletin, 85 (4), 845-857
HALL (1984): Nonverbal sex differences: Communication accuracy and expressive style. Baltimore & London: The John Hopkins University Press
HALPERN (2012): Sex Differences in Cognitive Abilities. 4th edition. New York: Psychology Press
HAUT & BARCH (2006): Sex influences on material-sensitive functional activation in working and episodic memory: Men and women are not all that different . NeuroImage , 32 , 411 – 422 .
HERDENER et al. (2010): Musical training induces functional plasticity in human hippocampus . Journal of Neuroscience , 30 , 1377 – 1384
HEYES (2010): Where do mirror neurons come from? In: Neuroscience and biobehavioral reviews. Band 34, Nummer 4, März 2010, S. 575–583,
HICKOK (2009): Eight problems for the mirror neuron theory of action understanding in monkeys and humans. In: Journal of cognitive neuroscience. Band 21, Nummer 7, Juli 2009, S. 1229–1243,
HICKOK (2015): Warum wir verstehen, was andere fühlen: Der Mythos der Spiegelneuronen. Übersetzt von Elsbeth Ranke. Carl Hanser Verlag, München 2015, 368 Seiten,
HINES (2004): Brain gender. Oxford and New York: Oxford University Press
HULSHOFF POL et al. (2006): Changing your sex changes your brain: Influences of testosteroneand estrogen on adult human brain structure. European Journal of Endocrinology,155(Suppl. 1), S107-S114.
IDIAKA et al. (2002): Age-related differnces in the medial temproal lobe responses to emotional faces as refeveald by fMRI. Hippocampus 12, 352 – 362
IM et al. (2008): Brain size and cortical structure in the adult human brain. Cerebral Cortex, 18, 2181 – 2191
INGALHALIKAR et al. (2014): Sex differences in the structural connectome of the human brain. PNAS January 14, 2014 111 (2) 823-828
INGALHALIKAR et al. (2014a): Reply to Joel and Tarrasch: On misreading and shooting the messenger PNAS February 11, 2014 111 (6) E638
JÄNKE et al. (2014): Brain size, sex, and the aging brain. Human Brain Mapping, 2014
JOEL (2011):Male or female? Brains are intersex. Frontiers in Integrative Neuroscience, 5 (57)
JOEL (2012): Generic-gondadal-genital sex (3G-sex) and the misconception of brain and gender, or, why 3G-males and 3G-females habe intersex brain and intersex gender. Biology of Sex Differences, 3 (27)
JOEL & TARRASCH (2014): On the mis-presentation and misinterpretation of gender-related data: The case of Ingalhalikar’s human connectome study. PNAS February 11, 2014 111 (6) E637
JOEL et al. (2015): Sex beyond the genitalia: The human brain mosaic. Proceesings of the National Academy of Sciences, 112 (50), 15468-15473
JOEL et al. (2016): Response to Del Guidice et al., and Rosenblatt: Do brains of females and males belong to two distinct populations? Proceedings of the National Academy of Sciences, 113 (14), E 1969 – 1970
JOST (1970): Hormonal Factors in the Sex Differentiation of the Mammalian Foetus. Philosophical Transactions of the Royal Society of London 259:119–130.
JORDAN-YOUNG (2010): Brain Storm The Flaws in the Science of Sex Differences. Cambridge: Harvard University Press
JURASKA et al. (1985): Sex differences in the dendritic branching of dentate granule cells following differential experience. Brain Res. 333, 73–80
JURASKA (1991): Sex differences in “cognitive” regions of the rat brain . Psychoneuroendocrinology , 16 , 105 – 19 .
KAISER et al. (2009): On sex/gender related similarities and differences in fMRI language research. Brain research Reviews, 61 (2): 49 – 59
KELLER & JUST (2016): Structural and functional neuroplasticity in human learning of spatial routes. Neuroimage Volume 125, 15 January 2016, Pages 256-266
KILLGORE & YURGELON-TODD (2001): Sex differences in the amygdala activation during the perception of facial affect. NeuroReport, 12 (11), 2543 – 2547
KINNUNEN et al. (2004): Differential brain activation in exclusively homosexual and heterosexual men produced by the selective serotonin reuptake inhibitor, fluoxetine. Brain Res. 2004 Oct 22;1024(1-2):251-4.
KOLATA (1983): Math genius may have hormonal basis. Science 222 (4630), 1312
KOROL et al. (2004): Role of estrogen in balancing contributions from multiple memory systems. Neurobiol. Learn. Mem. 82, 309–323
KOSOGONOV (2012): Why the Mirror Neurons Cannot Support Action Understanding. In: Neurophysiology. 44, 2012, S. 499-502,
KRUJIVER et al. (2000): Male-to-female transsexuals have female neuron numbers in a limbic nucleus. ournal of Clinical Endocrinology and Metabolism, 85, 2034-2041
KUTSCHERA, U. (2018): Das Gender-Paradoxon – Mann und Frau als evolvierte Menschentypen. LIT-Verlag
LALKLAR (2016): Vygotsky – a pioneering Soviet psychologist who derived his genius from Marxism
LAWRENCE (2003): Factors associated with satisfaction or regret following male-to-female sex reassignment surgery. Archives of Sexual Behavior, 32, 299-315.
LAWRENCE (2006): A Critique of the Brain-Sex Theory of Transsexualism
LENROOT & GIEDD (2010): Sex differences in the adolescent brain . Brain and Cognition , 72 , 48 – 55
LEONARD et al. (2008): Size matters: Cerebral volume influences sex differences in neuroanatomy. Cerebral Cortex 18 (12): 2920 – 2931
LEVAY ( 1991 ): A difference in hypothalamic structure between heterosexual and homosexual men . Science , 253 , 2034 – 1037
LEVITIN, K. 2009: One is not born a personality – A biographical history of Soviet Psychology. Erythros Press and Media
LIPPA (2005): Gender, Nature, and Nurture. 2nd ed. New York: Routledge
LIU et al. (2009): Evidence from intrinsic activity that asymmetry of the human brain is controlled by multiple factors . Proceedings of the National Academy of Sciences of the USA , 106 , 20499 – 20503
LUDERS, STEINMETZ & JANCKE (2008): Brain size and grey matter volume in the healthy human brain. NeuroReport, 13 (17), 2371 – 2374
LUDERS, THOMPSON & TOGA (2010): The development of the corpus callosum in the healthy brain . Journal of Neuroscience , 30 , 10985 – 10990 .
MACKEY et al. (2015): Neuroanatomical Correlates of the Income-Achievement Gap. Psychological Science Volume: 26 issue: 6, page(s): 925-933
MADEIRA & LIEBERMAN (1995): Sexual dimorphism in the mammalian limbic system. Prog. Neurobiol. 45, 275–333
MALL (1909): On several anatomical characters of the human brain, said to vary accorind to race and sex, with special reference to the weight of the frontal lobe. American Journal of Anatomy 9, 1 – 32
MARWHA, HALARI & ELIOT (2017): Meta-analysis reveals a lack of sexual dimorphism in human amygdala volume. NeuroImage Volume 147, 15 February 2017, Pages 282-294
MASCIE-TAYLOR (1993): How do social, biological, and genetic factors contribute to individual differences in cognitive abilities? In T. J. Bouchard Jr. & P. Propping (Eds.), Twins as a tool of behavioral genetics . Life sciences research report (pp. 53 – 65 ). Chichester, UK: John Wiley & Sons
MAY (2011): Experience-dependent structural plasticity in the adult human brain. Trends in Cognitive Sciences, 15(10), 475–482.
MCCARTHY et al (2015): Surprising origins of sex differences in the brain. Hormones and Behaviour, 76, 3-10
MCCLURE (2000): A meta-analytic review of sex differences in facial expression processing and their development in infants, children, and adolescents. Psychological Bulletin, 126 (3), 424-453
McDANIEL (2005): Big-brained people are smarter: A meta-analysis of the relationship between in vivo brain volume and intelligence. Intelligence. 2005; 33:337–346.
MCGLONE (1980): Sex differences in human brain asymmetry: A critical survey . Behavioral and Brain Sciences , 3 , 215 – 227
MEYER (2015): Adams Apfel und Evas Erbe. Wie die Gene unser Leben bestimmen und warum Frauen anders sind als Männer. München: Bertelsmann
MILLER (2008): Growing pains for fMRI. Science, 320 (5882), 1412 – 1414
MOORE (1995): Maternal Contributions to mammalian reproductive development and the divergence of male and female. Advances in the Study of Behavior, 24, 47-118
NIELSEN et al. (2013): An evaluation of the left-brain vs. Right-brain hypothesis with resting state functional connectivity magnetic resonance imaging, PLoS One 8 (8): e71275
NISHIZAWA et al. (1997): Differences between males and females in rates of serotonin synthesis in human brain. Proc. Natl Acad. Sci. USA 94, 5308–5313
NOBLE et al. (2015): Family income, parental education and brain structure in children and adolescents. Nature Neuroscience volume 18, pages 773–778
NOSTRO et al. (2017): Correlations Between Personality and Brain Structure: A Crucial Role of Gender. Cerebral Cortex, Volume 27, Issue 7, 1 July 2017, Pages 3698–3712
O’BOYLE (2005): Some current findings on brain characteristics of the mathematically gited adolescent. International Education Journal, 6 (2), 247 – 251
O’BOYLE et al. (2005): Mathematically gifted male adolescents activate a unique brain network during mental rotation. Cognitive Brain Resarch, 25 (2), 583-587
OBERMAN et al. (2005): EEG evidence for mirror neuron dysfunction in autism spectrum disorders. Cognitive Brain Reasearch, 24, 190-198
OHNISHI et al. (2004): The neural network of the mirror system and mentalizing in normally developed children: An fMRI study. NeuroReport, 15 (9), 1483 – 1487
OLIVEIRA (2009): Social behavior in context: hormonal modulation of behavioral plasticity and social competence. Integr. Comp. Biol. 49: 423-440
ORZHEKHOVSKIA (2005): Sex dimoprhism of neuron-glia correlations in the frontal areas of the human brain. Morfologiia, 127(1), 7 – 9
PACKARD et al. (1996): Posttraining intra-hippocampal estradiol injections enhance spatial memory in male rats: interaction with cholinergic systems. Behav. Neurosci. 110,626–632
PASCOLO et al. (2010): Critical review of the research leading to the mirror neuron paradigm – biomed 2010. In: Biomedical sciences instrumentation. Band 46, 2010, S. 422–427
PFANNKUCHE, BOUMA & GROOTHUIS (2008): Does testosterone affect lateralization of brain and behaviour? A meta-analysis in humans and other animal species Philos Trans R Soc Lond B Biol Sci. 2009 Apr 12; 364(1519): 929–942.
PINKER (2008): The Sexual Paradox. Men, Women and the Real Gender Gap. New York: Scribner/Simon& Schulster
RAINGRUBER (2001): Settling into and moving in a climate of care: Styles and patterns of interaction between nurse psychotherapist and clients. Perspectives in Psychiatric Care 37 (1), 15 – 27
REIS & CAROTHERS (2014): Black and White or Shades of Gray: Are Gender Differences Categorical or Dimensional? Current Directions in Psychological Science, 2014. 23(1): p. 19-26
RIDGWAY (2013): Illustrative effect sizes for sex differences
RIPPON (2016): „The trouble with girls?“. The Psychologist. British Psychological Society. 29 (12): 918–922.
RIPPON et al. (2014): Recommendations for sex/gender neuroimaging research. Frontiers in Human Neuroscience, 8, 1–13.
RITCHIE et al. (2018): Sex Differences in the Adult Human Brain: Evidence from 5216 UK Biobank Participants. Cerebral Cortex, Volume 28, Issue 8, 1 August 2018, Pages 2959–2975
ROBINSON et al. (1977): Monoamine metabolism in human brain. Arch. Gen. Psychiatry 34, 89–92
ROSE & ROSE (2001): Alas Poor Darwin – Arguments against Evolutionary Psychology. Vintage Verlag
RUIGROK et al. (2014): A meta-analysis of sex differences in human brain structure. Neurosci Biobehav Rev. 39(100): 34–50.
RUSHTON (1992): Corrections to a paper on race and sex differences in brain size and intelligence in Personality and Individual Differences 15(2):229-231
SATTERTHWAITE et al (2015): Linked Sex Differences in Cognition and Functional Connectivity in Youth Cerebral Cortex, Volume 25, Issue 9, 1 September 2015, Pages 2383–2394,
SAVIC, BERLUND & LINDSTRÖM (2005): Brain response to putative pheromones in homosexual men. Proc Natl Acad Sci U S A. 2005 May 17; 102(20): 7356–7361.
SAX (2006): Why gender matters: What parents and teachers need to know about the emerging science of sex differences. New York: Broadway Books
SCHNEIDER et al. (2000): Gender differences in regional cerebral activity during sdaness. Human Brain Mapping, 9, 226-238
SHORS (2016): A trip down memory lane about sex differences in the brain. Philosophical Transactions of the Royal Society B. doi: 10.1098/rstb.2015.0124.
SHORS et al. (2001): Sex differences and the opposite effects of stress on dendritic spine density in the male versus female hippocampus. Journal of Neuroscience, 21 (16), 6292 – 6297
SINGER et al. (2004): Empathy for pein involves the affecitve but not sensory components of pain. Science, 303 (1157), 1157 – 1162
SINGH & O’BOYLE (2004): Interhemispheric interaction during global-local processing in mathematically gifted adolescents, average-ability youth, and college studtentd. Neuropsychology, 18 (2): 371 – 377
SOMMER et al. (2004): Do women really have more bilateral language representation than men? A meta-analysis of functional imaging studies Brain, Volume 127, Issue 8, 1 August 2004, Pages 1845–1852,
SPELKE & GRACE (2007): Sex, math, and science . In S. J. Ceci & W. M. Williams (Eds.), Why aren’t more women in science: Top researchers debate the evidence (pp. 57 – 67 ). Washington, DC : American Psychological Association
TAN et al. (2016): The human hippocampus is not sexually-dimorphic: Meta-analysis of structural MRI volumes. NeuroImage Volume 124, Pages 350 – 366
TURNER & WEAVER (1985): Sexual dimorphism of glucocorticoid binding in rat brain. Brain Res. 343, 16–23
UDDIN et al. (2005): Self-face recognition activates a frontoparietal ‚mirro‘ network in the right hemisphere: An event-related fMRI study. NeuroImage, 25, 926-935
VAN ANDERS (2013): Beyond masculinity: testosterone, gender/sex, and human social behavior in a comparative context. Front. Neuroendocrinol. 34 (3) , pp. 198-210.
VAN ANDERS & WATSON (2006) Social neuroendocrinology: effects of social contexts and behaviors on sex steroids in humans. Hum. Nat. 17: 212-237
WAGNER et al. (2003): Valence, gender, and lateralization of functional brain anatomy in emotion: A meta-analysis of findings from neuroimaging. NeuroImage, 19 (3), 513 – 531
WALLEN (2005): Hormonal Influences on sexually differentiated behavoiur in nonhuman primates. Frontiers in Neuroendocrinology, 26, 7-26
WALLENTIN (2009): Putative sex differences in verbal abilities and language cortex: A critical review Brain and Language Volume 108, Issue 3, March 2009, Pages 175-183
WALSH et al. (2014): General and specific effects of early-life psychosocial adversities on adolescent grey matter volume. NeuroImage: Clinical Volume 4, 2014, Pages 308-318
WELCOME et al. (2009): Behavioral correlates of corpus callosum size: Anatomical/behavioral relationships vary across sex/handedness groups . Neuropsychologia , 47 , 2427 – 2435 .
WELLS (1989): Sigmund Freud. Kritik auf der Grundlage der Erkenntnisse von Iwan P. Pawlow. München: Verlag zur Förderung der wissenschaftlichen Weltanschauung
WOLLET & MAGUIRE (2011): Acquiring “the Knowledge” of London’s Layout Drives Structural Brain Changes. Current Biology, Vol 21 (24), S. 2109-2114
WOOD, HEITMILLER, ANDREASEN & NOPOULOS (2008): Morphology of the ventral frontal cortex: Relationship to femininity and social cognition. Cerebral Cortex 18: 534 – 540
WOOD, MURKO, NOPOULOS (2008): Ventral frontal cortex in children: Morphology, social cognition and femininity/masculinity. SCAN 3: 168 – 176
WOOLEY (2007): Acute effects of estrogen on neuronal physiology . Annual Review of Pharmacology and Toxicology , 47 , 657 – 680 .
WRAGA et al. (2006): Neural basis of stereotype-induced shifts in women’s mental rotation performance. Social Cognitive and Affective Neuroscience. 2, 12–19.
YOUNG & BALABAN (2006): Psychoneuroindoctrinology. Nature, 443, 634
ZHU et al. (1995): A sex difference in thehuman brain and its relation to transsexuality. Nature, 378, 68-70
