
Lebewesen können wie gesagt als Einzeller oder Mehrzeller vorkommen. Wenn viele Zellen zusammen einen Verbund eingehen, spricht man von Gewebe. Die meisten Tiere, Pilze und Pflanzen sind Vielzeller, bestehen also aus einem Verband mehrerer Zellen.
Gewebetypen bei Tierzellen
Ein Zellgewebe ist eine Ansammlung differenzierter Zellen einschließlich ihrer extrazellulären Matrix.
Die Zellen eines Gewebes besitzen ähnliche Funktionen und erfüllen gemeinsam die Aufgaben des Gewebes. Alle Anteile der meisten Vielzeller lassen sich einem Gewebe zuordnen, bzw. sie sind von einem Gewebetyp produziert worden. Hier soll erstmal eine kurze Auflistung erfolgen:
Die tierischen Gewebe (Abb. 1) sind:
- a) Epithelgewebe: Zellschichten, die alle inneren und äußeren Oberflächen bedecken. Es wird grob in Oberflächen- und Drüsenepithelien gegliedert.
- b) Binde- und Stützgewebe: Gewebe, das für strukturellen Zusammenhalt sorgt und Zwischenräume füllt (hierzu gehören auch Knochen, Knorpel und Fettgewebe) und im weitesten Sinne weitere spezialisierte Gewebe (z. B. das Blut) hervorbringt.
- c) Muskelgewebe: Zellen, die durch kontraktile Filamente für aktive Bewegung spezialisiert sind.
- d) Nervengewebe: Zellen, aus denen Gehirn, Rückenmark und periphere Nerven aufgebaut sind.
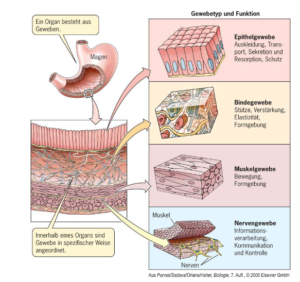
Abb. 1: tierische Gewebetypen
Oberflächenstrukturen und extrazelluläre Matrix
Die Oberfläche einer Zelle erfüllt verschiedene Aufgaben. Sie tritt in Kontakt mit anderen Zellen, grenzt sie aber gleichzeitig von anderen ab, regelt den Stofftransport in die Zelle und aus der Zelle hinaus und hat Rezeptoren für Signalmoleküle. Aufgrund ihrer verschiedenen Aufgaben sind Zelloberflächen außerordentlich vielgestaltig. Zu den Oberflächendifferenzierungen zählen so verschiedene Strukturen wie die Axone und Dendriten von Nervenzellen, Mikrovilli, Flagellen, Cilien und sonstige Zellfortsätze. Direkt mit der Cytoplasmamembran assoziiert ist die Glykokalyx. Darüber hinaus umgeben sich die Zellen mit extrazellulärem Material. Dieses wird von ihnen selbst sezerniert und bei den Tieren als extrazelluläre Matrix (Abb. 2), bei Pflanzen, Pilzen und Prokaryoten als Zellwand bezeichnet.
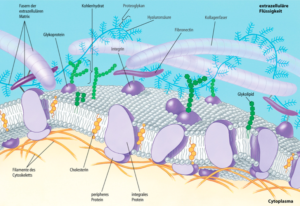
Abb. 2: extrazelluläre Matrix
Mit der Cytoplasmamembran sind eine Reihe von Glykoproteinen und Glykolipiden direkt oder indirekt assoziiert, deren teilweise sehr komplexen Oligosaccharid-Seitenketten ausschließlich auf der Außenseite exponiert sind. Der Besatz mit Glykoproteinen und Glykolipiden kann sehr dicht sein und einen regelrechten Zuckermantel um die Zellen bilden. Er wird als Glykokalyx bezeichnet. Der Zuckeranteil kann bis zu 10 % der Masse von Cytoplasmamembranen ausmachen. Die an der Oberfläche exponierten Strukturen sind für Zell-Zell-Erkennung, Zellkontakte und Signalwahrnehmung verantwortlich. Der Zellerkennung dienen beispielsweise Glykolipide, wie die Blutgruppenantigene des AB0-Systems oder die MHC-Komplexe, welche für die Gewebeverträglichkeit bei Organtransplantationen von entscheidender Bedeutung sind.
Die extrazelluläre Matrix (EZM) ist der Gewebeanteil, der zwischen den Zellen im sogenannten Interzellularraum bei tierischen Zellen liegt. Die extrazelluläre Matrix tierischer Vielzeller setzt sich aus diversen Komponenten zusammen, die in zwei große Gruppen eingeteilt werden: Grundsubstanz und Fasern. Die Fasern dienen der Formgebung und bestehen aus Kollagen oder Elastin, die in die Grundsubstanz eingebettet sind. Die Grundsubstanz besteht aus verschiedenen sog. Proteoglykankomplexen, Glucosaminoglykanen (wie Hyaluron) und/oder Glykoproteinen. Das Verhältnis der Grundsubstanz zum Faseranteil schwankt je nach Lokalisation ebenso wie der Anteil der extrazellulären Matrix am Gewebe insgesamt, bedingt durch dessen jeweilige Funktion.
Weitere wichtige Bestandteile der EZM sind Verbindungsmoleküle, die verschiedene Komponenten der EZM untereinander und diese mit der Zelloberfläche verbinden. Zwei der Wichtigsten sind Fibronektin und Laminin. Fibronektin enthält Bindungsstellen für Kollagen und Heparin. Durch alternatives Spleißen der mRNA entstehen verschiedene Fibronektin-Formen, die bei einer Reihe von biologischen Prozessen eine Rolle spielen. Dazu gehört die Blutgerinnung, Wundheilung oder die Wanderung der Neuralleistenzellen während der Embryonalentwicklung.
Laminin ist ein charakteristischer Bestandteil der Basallaminae und ähnlich wie Fibronektin enthält auch dieses Protein Bindungsstellen für verschiedene Moleküle wie Kollagen IV und Heparansulfat.
Die wichtigsten Rezeptoren an der Zelloberfläche für Komponenten der EZM sind die Integrine, die an verschiedene Liganden binden.
Zwischen den Zellen tierischer Gewebe gibt es eine Reihe von Zell-Zell-Verbindungen: Gap Junctions, Tight Junctions und Desmosomen.
Gap Junctions
Gap Junctions (Abb. 3) kommen vor allem in Muskel- und Nervengeweben tierischer Organismen vor, da dort eine schnelle Kommunikation erforderlich ist.
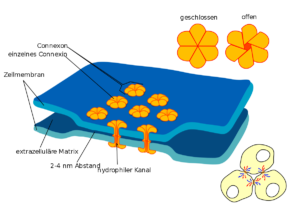
Abb. 3: Gap Junction
Es sind Proteinkanäle zwischen den Plasmamembranen zweier angrenzender tierischer Zellen. Sie ermöglichen somit eine direkte Verbindung des Cytoplasmas der beteiligten Zellen. Sie dienen zur Kommunikation zwischen Zellen, indem chemische Substanzen oder elektrische Signale weitergeleitet werden. sie werden etwa 2,7 nm groß.
Sie durchspannen die jeweiligen Plasmamembranen der benachbarten Zellen und den dazwischen liegenden Interzellularraum.
Das beteiligte Kanalprotein heißt Connexin. Sechs Connexine bilden nun jeweils eine ringförmige Struktur – das Connexon. Lagern sich jetzt jeweils zwei Connexone der benachbarten Plasmamembranen aneinander, bildet sich eine durchgehende Pore. Diese Pore entspricht einer Gap Junction.
Durch eine unterschiedliche Zusammensetzung der Connexine kann die Durchlässigkeit oder Leitfähigkeit der Kanäle jeweils variieren. Bei uns Menschen gibt es beispielsweise über 20 verschiedene Connexin-Gene. Der Durchmesser der Poren beträgt circa 1,5 nm, weshalb großen Molekülen wie Proteinen der Zutritt verwehrt bleibt.
Im Vergleich zu den anderen Kanalsystemen in der Zelle unterschieden sich die Gap Junctions vor allem darin, dass hier zwei Membranen anstatt einer durchzogen werden.
Die Hauptfunktion der Gap Junctions liegt in der Kommunikation zwischen benachbarten Zellen.
Eine Möglichkeit über Gap Junctions zu kommunizieren bietet die elektrische Kommunikation. Darunter versteht man eine schnelle Weiterleitung von elektrischen Signalen in Form von Ionenströmen. In Nervenzellen in der Netzhaut (Retina) am Auge oder im Herzen dienen sie als sogenannte elektrische Synapsen. Diese ermöglichen eine schnelle und gleichmäßige Ausbreitung von elektrischen Potentialen.
Neben der elektrischen Kommunikation kann auch eine chemische Kommunikation über Gap Junctions stattfinden. Signalmoleküle, sogenannte second messenger wie Calcium-Ionen (Ca2+), können die Gap Junctions durchqueren. Dadurch wird sichergestellt, dass diese Signalmoleküle in benachbarten Zellen in gleicher Konzentration vorhanden sind. Das ermöglicht eine koordinierte Reaktion der Zellen.
Außerdem können kleine Moleküle wie ATP, Glucose oder Aminosäuren durch die Kommunikationskanäle passieren. Sie sind für den Stoffwechsel in unseren Zellen sehr wichtig.
Gap Junctions sind nur in vielzelligen Tieren zu finden. Sie kommen vor allem dort vor, wo eine schnelle Kommunikation erforderlich ist, wie bei Muskel- und Nervenzellen. Zusätzlich unterstützen sie in Drüsen wie Leber und Bauchspeicheldrüse die Ausscheidung von Verdauungssekreten.
Desmosomen
Die Desmosomen sind in tierischen Zellen vertreten (Abb. 4). Man findet sie insbesondere in den Zellen des Herzmuskels und den Epithelzellen.
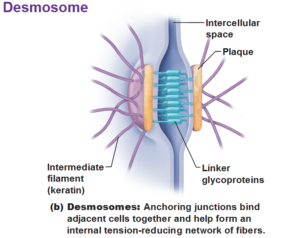
Abb. 4: Desmosomen
Ein Desmosom ist eine kugelförmige Struktur in der Zellmembran, vergleichbar mit Druckknöpfen.
Die wichtigste Funktion der Desmosomen ist die Herstellung einer Verbindung zwischen zwei Zellen, um dadurch den mechanischen Zusammenhalt zu erhöhen.
Die Zellstrukturen, welche eine Zelle nicht mit einer anderen Zelle, sondern mit dem Extrazellulärraum verbinden, werden als Hemidesmosomen bezeichnet.
Ein Desmosom setzt sich aus einem Netzwerk von Proteinen zusammen. All diese Proteine sind an der sogenannten Plaque befestigt. Man kann sich die Plaque als eine Art Verbindungsstelle im Desmosom vorstellen, die vor allem aus Ankerproteinen wie zum Beispiel Plaktoglobin und Plaktophilin besteht. Wie der Name schon sagt, dienen Ankerproteine als Verbindungsstücke für weitere Proteine.
An die Ankerproteine der Seite der Plaque, die zum Extrazellulärraum zeigt, sind die Cadherine wie zum Beispiel Desmoglein und Desmocollin befestigt. Diese länglichen Glykoproteine gehören zu den Adhäsionsproteinen. Ihre Aufgabe ist die Stabilisierung der Zell-Zell-Kontakte, indem sie sich mit den Cadherinen der anderen Zellen verbinden.
Die Seite der Plaque, die zum Zellinnenraum zeigt, enthält vor allem das Desmoplakin. Dieses spezielle Protein kommt nur in Desmosomen vor. Es bindet sich an die Ankerproteine und stellt eine Verbindung zu den Intermediärfilamenten des Cytoskeletts her. Diese Verbindung wird insbesondere in den Epithelzellen über sogenannte Fibrillenbündel aus den Proteinen Keratin oder Cytokeratin (Zytokeratin) erzeugt. Diese Fibrillenbündel aus Keratin werden auch als Tonofilamente bezeichnet.
Durch die Verknüpfung mit dem Cytoskelett können sich die Desmosomen an ihrem Ort an der Zelloberfläche stabilisieren.
Die Hauptaufgabe der Desmosomen ist die Verankerung der Zellen in Tieren untereinander (Zell-Zell-Kontakte).
Die Desmosomen stellen eine sehr enge Verbindung zwischen den Zellen her. Der Abstand der Plasmamembranen im Desmosom ist nur ungefähr 15-25 nm groß. Dadurch ist der gesamte Zellverbund gegenüber äußeren mechanischen Einwirkungen (Zug-/Scherkräfte) relativ stabil. Aus diesem Grund befinden sie sich insbesondere in Zellen mit hoher mechanischer Belastung (z.B. Herzmuskel).
Doch nicht nur die Stabilität des Zellverbundes gegen äußere Einflüsse kann durch Desmosomen erhöht werden. Indem sie eine Verbindung zwischen der Zellmembran und dem Cytoskelett beispielsweise über die Tonofilamente herstellen, können sie auch eine einzelne Zelle relativ gut in ihrer Form halten.
Die Hemidesmosomen sind eng mit den Desmosomen verwandt. Wie auch ein Desmosom, kommt ein Hemidesmosom sehr oft in Epithelzellen vor.
Wenn man ihren Gesamtaufbau in einem Elektronenmikroskop betrachtet, wird auffallen, dass Hemidesmosmen wie halbe Desmosomen aussehen. Sie enthalten ebenfalls eine Plaque, an die nun aber keine Desmogleine, sondern Kontaktproteine namens Integrine gebunden sind. Außerdem sind sie gleichermaßen intrazellulär mit den Intermediärfilamenten des Cytoskeletts über Keratinfilamente verknüpft.
Ihre Funktion besteht nun jedoch nicht mehr darin, zwei Zellmembranen ähnlicher Zellen zu verbinden. Hemidesmosomen sind in der Lage, die Zellmembran der Epithelzellen mit den extrazellulären Basallamina zu verbinden. Das ist die Proteinschicht, die an den basalen Teil der Epithelzelle anschließt. Diese Basallamina können sich über sogenannte Laminin Proteine, mit den Integrinen der Hemidesmosomen verbinden.
Tight Junctions
Tight Junctions kommen in tierischen Zellen vor allem im Darm, Magen und in der Blut-Hirn-Schranke vor. Sie sorgen für eine Verbindung zwischen Epithelzellen (Abb. 5).
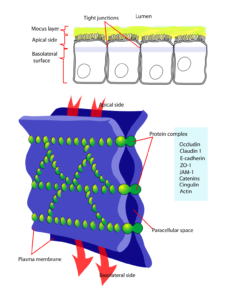
Abb. 5: Tight Junction
Tight Junctions kann man sich als eine Art reißverschlussartiges Proteinnetzwerk vorstellen, das sogenannte Verschlusskontakte bildet. Das bedeutet, dass kein Spalt (=kein Interzellularraum) zwischen den benachbarten Zellen übrigbleibt. Die Öffnungen sind also eng, was bereits der Name tight andeutet. Durch diese Abdichtung wird der Transport von Molekülen über die Epithelzellen kontrolliert. Es wird sichergestellt, dass keine Diffusion von gelösten Stoffen zwischen den Epithelzellen möglich ist.
Bei höheren Pflanzen ist hier übrigens analog dazu der Caspary-Streifen zu finden. Dieser sorgt in der Wurzel dafür, dass Schwermetallionen und andere Schadstoffe nicht in die Pflanze gelangen.
Tight Junctions sind aus einem gürtelartigen Netzwerk aus Membranproteinen um die Epithelzellen herum aufgebaut.
Das Netzwerk ergibt sich, indem spezielle Proteine in den Plasmamembranen der jeweiligen Epithelzellen aneinanderbinden. Diese Proteine sind in Strängen angeordnet und bilden eine Aneinanderreihung der einzelnen Kontaktpunkte.
Die wichtigsten daran beteiligten Membranproteine sind Occludin oder Proteine der Gruppe der Claudine. Je nach Zusammensetzung dieser Proteine ergeben sich unterschiedlich dichte Barrieren. Im Darm ist das Netzwerk beispielsweise loser aufgebaut, während es an der Blut-Hirn-Schranke einen dichteren Aufbau aufweist.
Tight Junctions haben neben der mechanischen Funktion, die darin besteht für Stabilität im Epithelverband zu sorgen, noch zwei weitere Funktionen: die Barrierefunktion und die Zaunfunktion.
Durch das komplexe Proteinnetzwerk der Tight Junctions entsteht eine Diffusionsbarriere, die eine Bewegung von gelösten Stoffen durch den Raum zwischen den Epithelzellen verhindert. Dadurch kann der Stoffaustausch auf zellulärer Ebene zwischen Körper und Umgebung kontrolliert werden. So muss beispielsweise jeder Stoff, bevor er vom Darm in den Körper gelangt, die Epithelzellen des Darms durchqueren. Auch in der Blasenwand sorgen Tight Junctions dafür, dass Urin nicht in die Bauchhöhle gelangt.
Außerdem besitzen Tight Junctions eine sogenannte Zaunfunktion. Darunter versteht man, dass die freie Bewegung der Membrankomponenten selbst verhindert wird. Das bedeutet, dass eine seitliche (=laterale) Diffusion der Membranproteine und Phospholipide gemäß dem Flüssig-Mosaik-Modell nicht möglich ist.
Dadurch wird gewährleistet, dass verschiedene funktionelle Bereiche mit unterschiedlicher Membranzusammensetzung in den Epithelzellen bestehen können. Die apikale Region (= Scheitelregion) unterscheidet sich beispielsweise von der basalen Seite (= Unterseite). Dies bezeichnet man als Zellpolarität.
Mikrovilli und Mikrofilamente
Zelloberflächen tierischer Zellen können auch unterschiedliche Ausbuchtungen enthalten. In den Tierzellen sind die wahrscheinlich wichtigsten Membranausbuchtungen die Mikrovilli. An ihrer Ausbildung sind vor allem Aktinfilamente beteiligt.
Die sogenannten Mikrofilamente oder auch Aktinfilamente sind Bestandteile des Cytoskeletts (Abb. 6). Sie sind mit einer Größe von nur etwa 6 nm die kleinsten der drei Filamente.
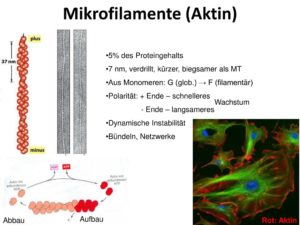
Abb. 6: Mikrofilamente
Wie der Name Akinfilament schon sagt, besteht es zum Großteil aus dem Strukturprotein namens Aktin, die die Monomere der Mikrofilamente bilden. Aktin ist eines der häufigsten Proteine in Eukaryoten. Sie entstehen durch Polymerisation aus einzelnen Molekülen des Strukturproteins Aktin.
Dabei bindet ein globuläres Aktin an ein ATP. Dieses Monomer (ATP-Aktin) kann sich nun mit weiteren Aktinmolekülen verbinden – polymerisieren, wobei ATP-Aktin unter Abspaltung (Hydrolyse) eines anorganischen Phosphatrestes zu ADP-Aktin wird. Die entstehende Kette von Aktinmonomeren bildet so die filamentöse Form der Aktinfilamente, auch F-Aktin genannt. Aktinfilamente sind aus zwei helikal miteinander verwundenen Aktineinzelfäden aufgebaut. Aktinfilamente weisen eine Polarität auf und haben ein schnell polymerisierendes sogenanntes (+)-Ende und ein langsam polymerisierendes (-)-Ende. ATP-Aktin bindet bevorzugt am (+)-Ende und das Filament wächst an diesem Ende. Am (–)-Ende läuft die Hydrolyse von ATP zu ADP schneller als die Anlagerung eines neuen ATP-Aktins ab, sodass ADP-Aktin dissoziiert und das Filament von dieser Seite verkürzt wird. Zahlreiche Begleitproteine steuern die Polymerisations- und Abbauvorgänge. Im Muskel werden die Filamente beispielsweise durch das Tropomyosin stabilisiert, das sich auf ganzer Länge an ein Filament anlegt. In Zellen außerhalb des Herzens und der Skelettmuskulatur wird Caldesmon gebildet.
Eine große Gruppe von Begleitproteinen, die auch als Aktin-bindende Proteine (ABP) bezeichnet werden, vernetzt Aktinfilamente untereinander und mit anderen Proteinen. Fimbrin, Villin, Filamin und Espin bilden Querverbindungen und so mechanisch steife Bündel (Abb. 7).
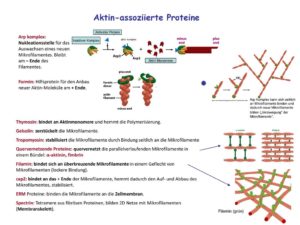
Abb. 7: Aktin-Assozierte Proteine
Aktin und Aktin-bindende Proteine sind an der Ausbildung der Mikrovilli (Abb. 8) beteiligt. Die Mikrovilli kommen vor allem in tierischen Epithelzellen vor.
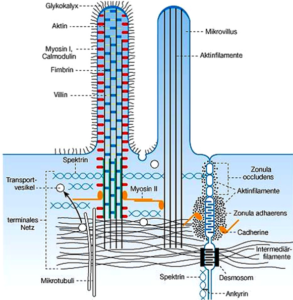
Abb. 8: Mikrovilli
Mikrovilli sind fadenförmige, leicht bewegliche Ausstülpungen der Zellmembran. Diese sind aus mehreren Bündeln von Aktinfilamenten aufgebaut. Ein Mikrovillus hat in etwa einen Durchmesser von 50 – 100 nm und ist 1 – 2 µm lang. Er besteht aus mehreren Bündeln von Aktinfilamenten. Die Bündel werden über die Proteine Fascin, Fimbrin und Villin in ihrer Form gehalten und stabilisiert.
Ihre Hauptfunktion besteht in der Oberflächenvergrößerung von Zellen. Durch diese größere Oberfläche können die Zellen Stoffe besser aufnehmen und abgeben. Diese ist vor allem bei den Epithelzellen wichtig, die körpereigene oder körperfremde Stoffe aufnehmen (= resorbierende Epithelien). Diese Zellen findet man vor allem in der Darmwand von Dünndarm und Dickdarm oder in den Nieren. Mit einer größeren Oberfläche können diese Stoffe schneller aufgenommen und abgegeben werden. Ohne die Mikrovilli würde der Dünndarm nur eine Oberfläche von etwa 4m2 besitzen. Mit ihnen kann sich die Fläche des Dünndarms auf bis zu 200m2 erstrecken.
Die Mikrovilli liegen nur auf der äußeren, der sogenannten apikalen Seite der Epithelzellen. Die innere, basale Seite enthält keine Ausstülpungen.
Um die Mikrovilli zusätzlich zu stabilisieren, ragen die Aktinfilamente mit dem Minus-Ende weiter in die Zelle hinein. Dort bilden sie das sogenannte Terminalgeflecht, das vor allem durch die Strukturproteine Tropomyosin und Myosin zusammengehalten wird. Das Strukturprotein Spektrin verankert die Mikrovilli weiterhin noch am Cytoskelett.
In ihrer Gesamtheit bilden die Mikrovilli den sogenannten Bürstensaum. Dieser Bürstensaum ist von einer Schicht aus Glykoproteinen und Glykolipiden namens Glykokalix überzogen, die die Zellmembran stabilisiert.
Mikrofilamente und Muskelgewebe
Eine weitere Aufgabe der Aktinfilamente findet man in Muskeln wieder (Abb. 9). Innerhalb der Muskelzellen liegen besonders viele Aktinfilamente. Diese tragen dazu bei, dass die Muskeln anspannen (Kontraktion) oder entspannen (Relaxation) können.
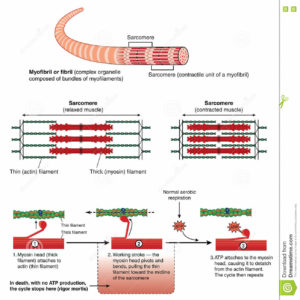
Abb. 9: Muskelgewebe
Die Muskelkontraktion ist ein biologischer Prozess, bei dem mechanische Kräfte im Muskelgewebe erzeugt werden. Im Falle der Skelettmuskeln werden diese Kräfte durch Sehnen auf die Knochen übertragen.
Die Kräfte entstehen durch Umwandlung von chemischer in mechanische Energie mittels des Aktin-Myosin-Komplexes in den einzelnen Muskelzellen, der wiederum seine chemische Energie aus der Hydrolyse von ATP bezieht.
Diese Bewegung wird durch Änderungen der chemischen Konfiguration und damit der Form der Myosin-Moleküle ermöglicht: Das Myosin besitzt kleine Fortsätze („Köpfe“), die ihren Winkel zum Rest des Moleküls („Schaft“) verändern können. Die Köpfe können wiederum an die Aktin-Filamente binden und diese in sogenannten „Ruderbewegungen“ verschieben. Im Ruhezustand (entspannter Muskel) ist das Aktinfilament mit so genannten Tropomyosinfäden umschlungen, die die Bindungsstellen der Myosinköpfchen an dem Aktinfilament bedecken. An das Myosin ist ATP gebunden, das Köpfchen befindet sich in einem 90-Grad-Winkel zum Schaft des Moleküls.
Ein Nervenimpuls bewirkt die Ausschüttung von Calcium (Ca2+). Das hat zwei Folgen: Zum einen aktiviert Ca2+ die Abspaltung von ATP in ADP + P am Myosinköpfchen, Als Cofaktor wird zusätzlich Mg2+ gebraucht. Das Calcium bindet zum anderen an Troponin, das an den Tropomyosinfäden angelagert ist, und verändert dabei deren Konfiguration so, dass die Bindungsstellen freigegeben werden und das Myosin an das Aktin binden kann.
Sobald das Myosin an das Aktin gebunden hat, wird das immer noch am Myosinköpfchen anliegende Pi und kurz danach auch das ADP freigesetzt. Dadurch wird die Verspannung des Myosins in mechanische Energie umgesetzt: Die Myosinköpfchen kippen in einem 45 Grad-Winkel zum Myosinfilament und ziehen dabei die Aktinfilamente von rechts und links zur Sarkomermitte.
Der Zyklus wird dadurch abgeschlossen, dass sich neues ATP an das Myosin anlagert. Dadurch löst sich das Myosinköpfchen vom Aktinfilament und die beiden Proteine befinden sich wieder im Ausgangszustand. Steht ATP nicht mehr zur Verfügung, können sich die Moleküle nicht mehr voneinander lösen und es kommt zur Totenstarre.
Pflanzengewebe
Pflanzliche Zellen verfügen über eine Zellwand, die aus Cellulose besteht. Einige Algen und viele Pilze haben anstelle von Cellulose Chitin in ihre Zellwand eingebaut.
Pflanzliche Gewebe lassen sich in zwei Gewebetypen unterscheiden: Bildungsgewebe (Abb. 10) und Dauergewebe (Abb. 11). Bildungsgewebe besteht aus teilungsfähigen Embryonalzellen (dem Meristem).
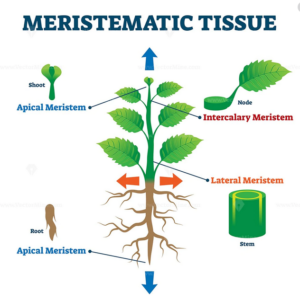
Die Zellen der Meristeme besitzen in der Regel dünne Zellwände mit wenig Cellulose und sind theoretisch unbegrenzt teilungsfähig. So gibt es Primärmeristeme wie das Protoderm, welches die Epidermiszellen bildet, während z. B. das Prokambium die Leitbündel bildet und das Grundmeristem das Markgewebe. Primärmeristeme bilden sich aus Apikalmeristemen und sind für das Primärwachstum der Pflanze zuständig Zum Sekundärmeristem gehört das Kambium, eine Gewebeschicht, die für das Dickenwachstum der Pflanzen zuständig ist. Bei Bäumen ist es die hohlzylinderförmige Wachstumsschicht zwischen der Splintholzzone und der Rinde, die auch Kambiumring genannt wird.
Wenn die Zellen nicht mehr teilungsfähig sind, handelt es sich um Dauergewebe (Abb. 11).
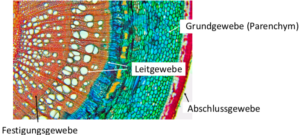
Abb. 11: Dauergewebe
Bei Pflanzen sind folgende Dauergewebe bekannt.
- a) Grundgewebe: hierzu zählt das Parenchym. Parenchymzellen sind dünnwandige Zellen des Grundgewebes, die den Großteil von nichtholzartigen (krautigen) Pflanzenstrukturen ausmachen und beispielsweise der Speicherung von Nährstoffen dienen.
- b) Festigungsgewebe: hier wird in Kollenchym und Sklerenchym unterschieden. Das Kollenchym besteht aus lebenden Zellen, mit nicht verholzten, dehnungsfähigen Zellwänden, das Sklerenchym aus toten Zellen mit verholzten Zellwänden.
- c) Das Abschlussgewebe welches in Epidermis und Periderm unterteilt wird. Die Epidermis ist das primäre Abschlussgewebe von Sprossachse und Blättern bei höheren Pflanzen. Das Periderm ist das sekundäre Abschlussgewebe der Sprossachse und das tertiäre Abschlussgewebe der Wurzel.
- d) Das Leitgewebe, welches in Xylem und Phloem geteilt wird. Das Xylem dient dem Wassertransport, das Phloem von Stoffwechselprodukten wie dem Zucker.
Diese vier Gewebetypen bilden bei höher entwickelten Pflanzen (also den sog. Kormophyten wozu die Farngewächse und Samenpflanzen gehören, nicht aber die Moose und Grünalgen) die typischen Organsysteme: Wurzel, Sprossachse und Blätter.
Plasmodesmen
Die Plasmodesmen sind kleine Plasmastränge in Pflanzenzellen (Abb. 12). Sie führen durch Aussparungen (= Tüpfel) in der Zellwand zu einer Nachbarzelle und schaffen so eine Verbindung, bei dem Stoffe transportiert werden können. Ein Plasmodesmos ist ein dünner Strang aus Plasma mit einem Durchmesser von etwa 30-50 nm.
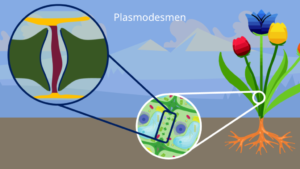
Abb. 12: Plasmodesmen
Innerhalb der Pflanzenzelle können sich auch mehrere Plasmodesmen befinden.
Da sie während der Zellteilung bestehen bleiben, sind sie in der Lage, die neu gebildeten Tochterzellen miteinander zu verbinden.
Abgesehen von den sogenannten Schließzellen enthalten wahrscheinlich alle Pflanzenzellen Plasmodesmen.
Schließzellen sind bohnenförmige Zellen mit Spaltöffnungen namens Stomata. Ihre Aufgabe ist insbesondere der Gasaustausch mit der umliegenden Luft durch Öffnen und Schließen der Stomata.
In seinem Inneren ist ein Plasmodesmos von einem Strang namens Desmotubulus durchzogen.
Seine Funktionen sind die Stabilisierung des Plasmodesmenkanal und die Verbindung zum Endoplasmatischen Retikulum und den Zellmembranen anderer Zellen. Insbesondere er ist in der Lage, Lipide, Proteine, RNA und weitere Moleküle zur anderen Zelle zu transportieren.
Es gibt drei Möglichkeiten, wie der Desmotubulus die Stoffe transportieren kann:
Zum einen kann er sie entlang seiner Membran bewegen. Zum anderen kann er den Stofftransport durch den zytoplasmatischen Bereich der Plasmodesmen steuern. Eine dritte Möglichkeit besteht darin, die Stoffe durch seinen Innenraum (= Lumen) zu transportieren.
Vor allem bei Letzterem ist er in der Lage, die Größe der weitergeleiteten Stoffe durch seinen Durchmesser zu begrenzen.
Für den Stofftransport innerhalb der Pflanzenzelle existieren zwei unterschiedliche Wege namens Apoplast und Symplast (Abb. 13). Diese beiden Transportwege werden durch die Plasmodesmen jeweils voneinander abgetrennt.
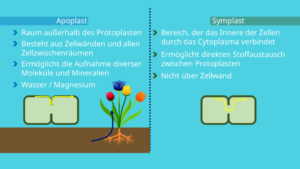
Abb. 13: Apoplast und Symplast
Der Apoplast bezeichnet den Raum außerhalb des Protoplasten. Dieser Raum besteht aus den Zellwänden und allen Zellzwischenräumen.
Über den Apoplasten ist die Zelle in der Lage, verschiedenste Moleküle oder Mineralien aufzunehmen. Das können zum Beispiel Wasser oder Magnesium sein. Beides erhält die Pflanze vom Boden. Diese Stoffe werden dann über die Zellwände verbreitet.
Der zweite Transportweg geht über den sogenannten Symplasten. Er bezeichnet den Bereich, der das Innere der Zellen durch das Cytoplasma miteinander verbindet.
Über den Symplast können Moleküle und Mineralien direkt zwischen den jeweiligen Protoplasen weitergegeben werden. Diese Weitergabe erfolgt hier nicht über die Zellwand, sondern direkt über die Plasmodesmen.
