

Fossilien sind Zeugen des Lebens der Vergangenheit und belegen, dass sich das Leben auf unserer Erde verändert. Wenn aber die Evolutionsbiologie davon ausgeht, dass alle Lebewesen auf einen gemeinsamen Vorfahren zurückgehen, dann muss es zwischen verschiedenen Gruppen von Lebewesen Übergänge geben. Wenn es um Übergangsformen geht, so kommen seitens der Kreationisten viele Vorurteile und sachliche Unkenntnisse, wenn nicht sogar argumentative Taschenspielertricks und Manipulation. In diesem Beitrag befassen wir uns daher mit der Frage, was grundsätzlich unter einer Übergangsform zu verstehen ist.
Lücken im Fossilbericht?
Text als pfd
Die Argumentation der Kreationisten lautet wie folgt: Wenn die Evolutionstheorie fordert, dass zwischen taxonomischen Gruppen Übergangsformen existieren, so müsste es Millionen und Abermillionen von Übergangsformen geben. Doch diese gäbe es angeblich nicht. Die Lücken im Fossilbericht bestätigen angeblich, dass die Lebewesen „fertig“ ausgebildet werden. Sie erlauben zwar gewisse sog. mikroevolutionäre Änderungen innerhalb der von Kreationisten willkürlich postulierten „Grundtypen“ (englisch: kinds), doch zwischen diesen Grundtypen gäbe es keine evolutionären Übergangsformen. Zum Thema „Grundtypen“ habe ich schon einen Beitrag verfasst, weswegen ich hier nicht noch einmal darauf eingehen werde. „Promiente“ Übergangsformen, wie z. B. Archaeopteryx, welcher den Übergang zwischen Raubdinosauriern und Vögeln belegt, werden dann nicht als Übergangsformen anerkannt, sondern – in diesem Fall – als „fertiger Vogel“.
Es fällt bei genauerem Hinsehen jedoch auf, dass Kreationisten ein Verständnis dafür fehlt, was eine Übergangsform tatsächlich darstellt. Nicht wenige, besonders radikale Kreationisten, kommen mit diversen Phantasietieren auf.
Wir wissen z. B., dass Vögel zusammen mit Dinosauriern und Krokodilen die Gruppe der Archosaurier bilden. D. h. all diese Gruppen gehen auf einen gemeinsamen Vorfahren zurück. Wie versuchen besonders skurrile Kreationisten dies ins Lächerliche zu ziehen? Sie behaupten, wenn Evolution stimme und Vögel und Krokodile näher miteinander verwandt seien, müsse es sowas wie eine Ente mit Krokodilkopf geben … eine sog. Croco-Duck (Abb. 1). Es werden also verschiedene Körperteile heute lebender Tiere willkürlich zusammengeführt und das soll dann die Übergangsform sein. Doch so funktioniert weder Evolution, noch werden Übergangsformen so verstanden.

Abb. 1: Die Kroko-Duck, welche den evolutionären Übergang zwischen Dinosauriern und Vögel ins Lächerlich ziehen soll.
Was also sind nun evolutionäre Übergangsformen?
Abstammung mit Modifikation in Stammbäumen
Wenn wir Lebewesen systematisch einordnen untersuchen wir ihre apomorphen Merkmale, die diese Lebewesen kennzeichnen und in ein hierarchisch verschachteltes System gemeinsamer Merkmale einordnen.
Jedes Lebewesen ist eine leicht modifizierte Version seiner Population.
Durch immer weiter fortlaufende Artbildungsprozesse, entstehen nicht nur neue Arten, sondern einige Gruppen der Arten ähneln sich stärker als anderen Gruppen, wodurch man sie in immer höhere Kladen zusammenfasst.
Wenn evolutionäre Linien sich weiter aufspalten, werden sie zunehmend genetisch verschieden. So können sich die entfernten Nachkommen der Ursprungsart so stark voneinander unterscheiden, dass es Sinn macht, sie in höhere taxonomische Kladen zu gruppieren. Gleichzeitig sind sie aber immer noch Teil der Abstammungslinie aus der sie hervorgegangen sind.
Wenn sich neue Arten im Stammbaum von ihren Vorläuferarten abspalten, sind sie immer noch Teil dieser Verwandtschaft und damit immer noch Teil der höheren Ränge der Verwandtschaft.
Pedantisch gesehen macht also ein Übergang zwischen Dinosauriern und Vögeln keinen Sinn, da Vögel eine Untergruppe der Dinosaurier sind. Genauso wenig stammt der Mensch vom Affen ab, wir sind nämlich Affen, genauso wie wir Säugetiere, Amnioten, Tetrapoden, Wirbeltiere etc. sind.
Aber wie kommt man überhaupt dazu, Enten und Krokodile, oder allgemein: Vögel und Reptilien, in eine verwandtschaftliche Beziehung zu stellen? Auf den ersten Blick wirkt das tatsächlich merkwürdig.
Vögel sind ganz klar zu erkennen, denn alle haben Federn, legen Eier, haben einen Schnabel und ihre Vordergliedmaßen sind zu Flügeln umgewandelt. Auch Reptilien sind für viele recht einfach zu erkennen: sie haben Schuppen und sind wechselwarm. Die Unterschiede zwischen Reptilien und Vögeln fallen also jedem auf und in diesem Sinn ist es nur logisch sie in zwei verschiedene Klassen einzuteilen, die auf den ersten Blick nichts miteinander zu tun haben.
Es gibt aber auch eine Reihe auffälliger Gemeinsamkeiten.
Beide Gruppen legen amniotische Eier: hartschalige Eier, bei denen die Embryonen im Ei mit mehreren Häuten umgeben sind, wodurch eine Entwicklung unabhängig vom Wasser ermöglicht wird. Weiterhin haben Vögel auch Merkmale, die man sonst bei Reptilien kennt: u. a. Schuppen (an den Beinen) oder Zehen mit Krallen – gleichzeitig lassen sich Federn morphologisch von Schuppen ableiten.
Forschern ist schon länger aufgefallen, dass Vögel einige Reptilienmerkmale haben und besonders die Krokodile mehr mit den Vögeln verwandt zu sein scheinen, als mit anderen Reptiliengruppen – dies zeigt sich z. B. am Bau des Herzens, Details in der Schädelanatomie oder genetischer Merkmale.
Und was passiert, wenn wir die evolutionäre Vergangenheit berücksichtigen und somit ausgestorbene Gruppen mit einbeziehen? Hier Fallen weitere Gemeinsamkeiten zwischen einigen Reptilien, in diesem Fall den Dinosauriern und Vögeln auf. Es ist bekannt, dass einige Dinosaurier, vor allem die Raubdinosaurier, wie Microraptor, Deinonychus oder Archaeopteryx, eine Reihe von Merkmalen besitzen, die typisch für Vögel sind. Sie haben z. B. Federn oder hohle Knochen. Durch solche Merkmalskombinaten verschwimmen die Grenzen zwischen den einzelnen Taxa zunehmend.
Vögel und Krokodile sind also nicht durch eine einzige Artaufspaltung voneinander getrennt. Den ersten Artbildungsprozessen folgten viele weitere, bei dem die Merkmale der einzelnen Kladen sich weiter ausbildeten. Zwei heute lebende Tierarten zu nehmen und sie irgendwie zusammenzufügen ist nicht das, was unter evolutionären Übergängen zu verstehen ist.
Eine gute Analogie lässt sich durch die Entwicklung der Sprachen demonstrieren (Abb. 2): Vergleicht man Spanisch und Französisch miteinander, wird auffallen, dass es zwischen beiden Sprachen Gemeinsamkeiten gibt. Das ist kein Zufall, denn spanisch und französisch stammen, neben anderen Sprachen, vom Lateinischen ab. Jedoch können wir ziemlich sicher sein, dass zwei lateinisch sprechende Eltern nicht ein Kind auf die Welt brachten, dass Französisch oder Spanisch sprach. Wie bei sich evolvierenden Populationen, entwickelten sich die Sprachen über Generationen und Zeiträume. Kleine Änderungen in der Aussprache, kleine grammatikalische Änderungen oder Wortneuschöpfungen über mehrere Generationen in isolierten Populationen sorgten dafür, dass sich aus der ursprünglichen lateinischen Sprache über diverse Dialekte französisch, spanisch, portugiesisch und italienisch entwickelten.
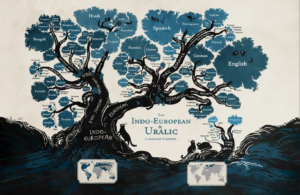
Abb. 2: Die Evolution der Sprache als Analogie zum evolutionären Stammbaum.
Die Suche nach den Vorfahren
Manche Kreationisten kommen dann jedoch mit einem anderen „Argument“: Sie sagen: ja mag sein, dass Archaeopteryx Merkmale von Dinosauriern und Vögeln gemeinsam hat, aber er ist nicht der Vorfahre aller Vögel, aufgrund diverser Details in der Anatomie.
Findet man Übergangsformen heißt das aber nicht automatisch, dass diese die direkten Vorfahren einer Gruppe sein müssen, sondern vielleicht Seitenäste in der Entwicklung. Die Erklärung dafür ist ganz einfach.
Über 99% aller Lebewesen sind im Verlauf der Erdgeschichte ausgestorben und nur ein winziger Bruchteil davon wird zum Fossil. Das heißt nur ein Bruchteil aller Arten wird überhaupt zum Fossil und es wird nicht jedes Fossil entdeckt. Folgerichtig, wenn nicht alle Arten zum Fossil werden und wir nicht alle Fossilien entdecken, ist es sehr vage zu behaupten Fossil X ist der Vorfahre der heute lebenden Art Y.
Wir können hierzu auch unseren eigenen Familienstammbaum als Vergleich nehmen. Wir sind das Produkt unserer Eltern. Aber unsere Eltern haben meist Geschwister, die ebenso Kinder haben. Wiederum kommen noch die Großeltern hinzu, die ebenfalls Geschwister haben usw. usf. Wenn jemand (z. B. ein Ahnenforscher) meinen Familienstammbaum untersuchen will, aber nur meine Groß-Tante ausfindig machen kann, so ist es offensichtlich, dass sie nicht meine Mutter sein kann. Dennoch ist nicht abzustreiten, dass sie mit mir verwandt ist. Und nur weil man meine Eltern nicht findet, heißt es nicht automatisch, dass ich durch einen göttlichen Akt geschaffen wurde. Genauso verhält es sich mit dem Fossilfund. Wenn wir Fossilien finden und sie verwandtschaftlich in die Nähe rezenter Arten rücken, sagen wir damit nicht, dass wir genau den Vorfahren entdeckt haben, der direkt zur untersuchten heute lebenden Art führte. Wir sagen lediglich, dass dieses Fossil eine Reihe von Merkmalen mit heute lebenden Arten hat, die auf eine gemeinsame Abstammung hindeuten. Es ist, wenn überhaupt, dann ein “Vorfahre” im weitesten Sinne, sodass wir z. B. sagen können, Vögel stammen von einer Gruppe von Raubdinosauriern ab und Archaeopteryx ist dafür ein Beispiel. Wir sagen aber nicht, die Art Archaeopteryx ist der Vorfahre aller heute lebenden Vögel.
Mosaikevolution
Die mosaikartige Evolution von Merkmalen bei Lebewesen wird von Kreationisten ebenfalls gerne missverstanden.
Beispiel Archaeopteryx
Schauen wir uns hier Archaeopteryx noch einmal an:
Archaeopteryx ist eine Mosaikform aus abgeleiteten und ursprünglichen Merkmalen (Abb. 3). Kreationisten sagen Archaeopteryx sei nur ein richtiger Vogel und keine Übergangsform, weil er typische Vogelmerkmale, wie etwa ein Federkleid, ein zum Gabelbein verwachsenes Schlüsselbein, ein nach hinten gerichtetes Schambein und Flügel hat.
Dass der Urvogel aber noch nicht alle Merkmale der modernen Vögel besaß, wird von Kreationisten zumeist ignoriert: Es fehlen ihm noch der für heutige Vögel typische Hornschnabel, das Pygostyl (die verschmolzenen, letzten Schwanzwirbel), die verwachsenen Finger, der einknöchrige Mittelfinger sowie das für Vögel typische knöcherne Brustbein. Darüber hinaus kamen ihm „Reptilienmerkmale“ zu, die wiederum den modernen Vögeln fehlen, wie z. B. Bauchrippen, ein bezahnter Kiefer, ein langer, knöcherner Schwanz sowie drei Fingerklauen an den Flügeln.
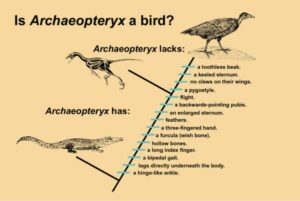
Abb. 3: Archaeopteryx als Bindeglied zwischen Reptilien und Vögeln.
Inzwischen hat sich die kreationistische Desinformation bezüglich Archaeopteryx dadurch erledigt, dass so viele in unterschiedlichem Ausmaß vogelähnliche, kleine Raubsaurier (Theropoda) fossil bekannt sind, dass es an dem Übergangsfeld von den Dinosauriern zu den Vögeln keinen vernünftigen Zweifel mehr geben kann (Abb. 4).

Abb. 4: befiederte Dinosaurier
Beispiel Tiktaalik
Dies lässt sich auch bei anderen berühmten Übergangsformen zeigen, die den Übergang an das Landleben bei Wirbeltieren dokumentieren. Landwirbeltiere werden gemein hin als Tetrapoden (Vierfüßer) bezeichnet.
So weisen die Gattungen Eusthenopteron und Panderichthys noch relativ wenig Tetrapoden-Merkmale auf, während sich die ursprünglichen Tetrapoden Acanthostega und Ichthyostega morphologisch bereits mehr oder weniger deutlich von ihnen unterscheiden (Abb. 5).
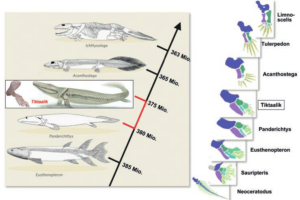
Abb. 5: Evolution der Tetraopden
Tiktaalik roseae (Abb. 5) verkörpert eine Zwischenform, die genau in diese morphologische Lücke fällt: Das Tier mit dem krokodilähnlichen Schädel steht aufgrund seines einzigartigen Merkmalsmosaiks im phylogenetischen System „zwischen“ den Raubfischen Panderichthys und den ältesten Tetrapoden Acanthostega und Ichthyostega (AHLBERG & CLACK 2006, S. 748;).
Das Tier lebte im Ober-Devon vor 380 Millionen Jahren und verfügt über Flossen, die den Beginn der Entstehung der Gliedmaßen dokumentieren. So besitzt die Extremität Oberarmknochen, Elle, Speiche und Handwurzelknochen, weist aber noch keine eindeutigen Finger auf (DAESCHLER et al. 2006; SHUBIN et al. 2006). Zudem zeigt das verkürzte Schädeldach, das Fehlen von Kiemendeckeln, die Schultergürtel und viele andere Merkmale Anklänge an die Tetrapoden, während andere Merkmale, wie die Flossenstrahlen, „fischartig“ geblieben sind.
Tiktaalik passt nicht nur aufgrund seines Körperbaus sehr gut in das zu erwartende Übergangsfeld zwischen den Devonischen Raubfischen und ursprünglichen Vierbeinern. Auch stratigraphisch fällt Tiktaalik exakt in die evolutionstheoretisch vorhergesagte Formation des untersten Ober-Devons (AHLBERG & CLACK 2006, S. 748). Tiktaalik ist damit etwa 3 Mio. Jahre jünger als Panderichthys und ungefähr 20 Mio. Jahre älter als die auf 365 Mio. Jahre datierten Überreste der Urtetrapoden Acanthostega und Ichthyostega.
Durch Tiktaalik wird nicht nur erneut die Vorhersagekraft der Evolutionstheorie in Bezug auf die Existenz von Zwischenformen bestätigt, sondern auch die Deszendenztheorie als wesentliches Teilstück der Evolutionstheorie massiv gestützt: Von den ältesten Fossilschichten aus betrachtet nähern sich die Lebewesen in stufenweiser Abänderung den heutigen Formen.
Auf der kreationistischen Homepage Genesisnet erschien 2006 eine interessante Stellungnahme von Reinhard JUNKER, einem führenden Kreationisten im deutschsprachigen Raum. Dieser erkennt zunächst an, dass „Tiktaalik … ein Merkmalsmosaik [besitzt], das gut in einen Übergangsbereich zwischen Fischen und Vierbeinern passt“ (JUNKER 2006). Dieses Zugeständnis ist im Grundsatz erfreulich, da es sich wohltuend von der „Schwarz-Weiß-Argumentation“ anderer Evolutionsgegner abhebt und sich durch etwas mehr Fairness in der Auseinandersetzung auszeichnet.
Gleichwohl schreibt Junker:
„Dass Merkmale von Tiktaalik besonders hervorgehoben werden, die als ‚Übergangsmerkmale‘ gedeutet werden können, ist legitim. Dennoch zeigt die Brustflosse insgesamt doch eher einen fischartigen Charakter (Ahlberg & Clack 2006, 748). Ein Vergleich mit anderen Formen aus dem Übergangsbereich Fische – Vierbeiner macht dies deutlich.“
Was mit dieser Aussage erreicht werden soll, bleibt unklar, denn die Frage, ob die Brustflosse von Tiktaalik eher tetrapodenartig oder „insgesamt doch eher fischartig“ war, ist in diesem Kontext irrelevant. Aus evolutionärer Perspektive ist nur wichtig, dass die Brustflossen von Tiktaalik in morphologischer Hinsicht zwischen denen devonischer Raubfische (wie Panderichthys) und den Extremitäten primitiver Tetrapoden (wie Ichthyostega) stehen (vgl. SHUBIN et al. 2006, AHLBERG & CLACK 2006).
Weiterhin schreibt Junker (2006):
„Die Unterschiede zwischen Tiktaalik und gefingerten Gattungen wie Acanthostega sind erheblich … Ahlberg & Clack (2006, 748) weisen darauf hin, dass der Erwerb von Fingern, von Tiktaalik ausgehend, eine erhebliche Umorganisation (‚developmental repatterning‘) erfordern würde. Der achtfingrige oberdevonische Tetrapode Acanthostega … war höchstwahrscheinlich ausschließlich wasserlebend; seine Extremitäten waren relativ unbeweglich. Insgesamt eignet sich diese Gattung daher nicht als vermittelnde Form zwischen Tiktaalik und landlebenden Tetrapoden.“
„Tiktaalik hatte deutlich andere ‚Übergangsmerkmale‘ auf dem Weg zum Landleben als Acanthostega. Der Weg aufs Land konnte nicht über beide Formen zugleich führen, es sei denn, er wurde mindestens zweimal unabhängig durchlaufen, womit dann aber die Vierbeinigkeit nicht mehr als Schlüsselmerkmal (d. h. als Hinweis auf gemeinsame Vorfahren) gelten könnte, sondern konvergent entstanden wäre.“
„Schon länger ist klar, dass auch das berühmte Ichthyostega …, deutlich von einer vermittelnden Position entfernt ist … Nicht nur der Bau der Flossen, sondern auch der Schädelbau passt insgesamt nicht in eine evolutive Reihe von tetrapodenähnlichen Fischen hin zu frühen Tetrapoden. Auch in dieser Hinsicht würde sich Acanthostega auf einer anderen ‚Schiene‘ bewegen, wenn man die relevanten fossilen Gattungen in evolutionäre Linien einfügen wollte“.
Kein Evolutionsbiologe, der konsequent auf dem Boden der phylogenetischen Systematik argumentiert, hat je gefordert, dass Panderichthys, Tiktaalik, Acanthostega und Ichthyostega auf „einer Schiene“ von „tetrapodenähnlichen Fischen hin zu frühen Tetrapoden“ überleiten müssten; dies wäre aus kladistischer Sicht geradezu widersinnig.
Daraus folgt auch nicht, dass die Vierbeinigkeit nicht mehr als ein von einem gemeinsamen Vorfahren der Tetrapoden erworbenes (Schlüssel-) Merkmal gelten könne. Die von SHUBIN et al. (2006, a.a.O.) diskutierten Homoplasien bedeuten zunächst nur, dass sich die Extremitäten in einigen Entwicklungslinien unabhängig voneinander ähnlich spezialisiert haben.
Die Aussage, die bekannten Fossilien seien nicht „intermediär“, lässt darauf schließen, dass Kreationisten aus evolutionärer Sicht geradezu eine harmonische, kontinuierliche Veränderung der einzelnen Merkmale erwarten.
Nach Ansicht der Evolutionsgegner dürfe eine „evolutionäre Umwandlung“ zumindest nicht über „spezielle Merkmale“ führen, denn einzigartige Strukturen und Verhaltensweisen können dazu führen, dass die Evolution in eine Sackgasse gerate, da spezialisierte Lebensweisen einer weiteren Entwicklung im Wege stünden.
Solch eine Argumentation würde nur Sinn ergeben, wenn Fachleute von einer Übergangsform erwarten würden, dass sie hinsichtlich aller Merkmale genau zwischen den vermittelnden Taxa stehen müsse.
Warum dies widersinnig ist, erklärte bereits der berühmte Evolutionstheoretiker Ernst MAYR (1967, 465–467):3)
„“Beim Wechsel in eine neue adaptive Zone steht eine Struktur oder ein Strukturkomplex unter besonders strengem Selektionsdruck […] Daraufhin evoluiert diese Struktur oder dieser Komplex besonders schnell, während andere zurückbleiben. Das Resultat ist nicht ein stetiger und harmonischer Wandel aller Teile des ‚Typus’, wie die idealistische Biologie sich das einbildet, sondern weit mehr eine Mosaikevolution. Jeder evolutive Typus ist ein Mosaik primitiver und fortschrittlicher Merkmale, von allgemeinen und spezialisierten Zügen.“
Neue Arten entstehen häufig durch die Aufspaltung von Populationen (Kladogenese), wenn auf die Teilpopulationen unterschiedlichen Selektionsfaktoren einwirken, oder wenn neue Adaptationen einer Population über längere Zeit durch Evolution entstehen.
In beiden Fällen ist ein mosaikartiges Nebeneinander von primitiven und fortschrittlichen Merkmalen der Übergangsformen (im Fachjargon Heterobathmie genannt) zu erwarten und tritt nahezu zwingend ein. D.h. man erkennt eine Übergangsform gerade daran, dass sie ein Mosaik aus abgeleiteten und ursprünglichen Merkmalen zeigt. Aus Sicht der Evolutionsbiologie ist das Auftreten von Heterobathmien völlig trivial (vgl. MAHNER 1986; FUTUYMA 1990; NEUKAMM & KUTSCHERA 2006).
Die Diskrepanz zwischen der modernen Auffassung über den Modus der Evolution und die veralteten Vorstellungen der Kreationisten hatte Mahner (1986, S. 62) mit folgenden Worten hingewiesen:
„Ein Grund kreationistischer Fehlinterpretation von ‚connecting link’ ist deren unterschiedlicher Auffassung im Gegensatz zu den Evolutionsbiologen. Während für diese das ‚connecting link’ ein Merkmalsmosaik darstellt, das durch die unterschiedliche Evolutionsgeschwindigkeit verschiedener Merkmale zustande kommt (Heterobathmie) und aus dem Nebeneinander von ursprünglichen und abgeleiteten Merkmalen besteht, verlangen die Kreationisten geradezu ein Morphenkontinuum zwischen den Fossilien (…) Präsentiert man ihnen eine Zwischenform, so wollen sie zwischen dieser und der Ahnenform eine weitere Zwischenform und so fort (…) Nicht nur solche Aussagen, sondern auch die fast ausschließliche Rede von der ‚Umwandlung’ einer Art in eine andere zeigen, dass die Kenntnisse der Kreationisten über Artbildung nicht zeitgemäß sind.“
Leiter, Büsche & Missing Links
Unabhängig von der Unwissenheit der Kreationisten zum Thema Evolution und Übergangsformen gibt es jedoch ein weiteres populäres Missverständnis bezüglich der Evolutionsdarstellungen.
Traditionell wird gerne von einer „Leiter“ der Evolution gesprochen, bei der sich „primitive“ Arten in „höher entwickelte“ verwandeln. Man spricht auch gerne über die große Seinskette, die durch Bindeglieder miteinander verbunden wird. Sind diese Bindeglieder nicht bekannt, spricht man gerne vom „missing link“.
Die berühmteste Darstellung der Vorstellung der Evolution als Leiter ist die Entwicklungsreihe vom Affen zum Menschen (Abb. 6). Als Übergänge sieht man dann Vertreter der Urmenschen wie Homo erectus und den Neandertaler. Diese Darstellung erweckt den Anschein, als ob es eine Evolutionsreihe gab, die ohne weitere Abspaltungen vom primitiven Affen hin zum modernen Menschen führte.

Abb. 6: Die Evolution der Menschen als Beispiel für eine Stufenleiter
Die Bedeutung des missing links sollte also besagen, dass in der darwinschen Theorie eine entscheidende Verbindung zwischen Menschen und anderen Tieren, allen voran den Affen, fehlte.
Diese Vorstellung ist der Mythos der großen Seinskette, wonach alle Dinge im Universum auf einer Leiter stehen. Ganz oben befindet sich Gott, dann die Engel, dann die Menschen, dann die Tiere, dann die Pflanzen und ganz unten schließlich die Steine und weitere unbelebte Dinge. Und natürlich stehen in dieser großen Seinskette nicht alle Menschen auf einer Stufe, denn Männer stehen angeblich höher als Frauen und die Weißen über den Schwarzen.
Dieser Mythos der großen Seinskette fand auch Einklang in die Vorstellungen zur Evolution im viktorianischen Zeitalter und hält sich bis heute hartnäckig in den Vorstellungen unserer Kultur. So hätten sich „niedere“ Tiere, sagen wir mal der Regenwurm, zu „höheren“ Tieren, sagen wir z. B. den Affen entwickelt. Und aus den Affen entwickelte sich der „höher entwickelte“ Mensch. Das Problem dieser Vorstellung ist, dass sie eine zutiefst falsche und unevolutionäre ist. Denn mit welchem Recht können wir aus evolutionsbiologischer Sicht behaupten der Wurm ist primitiver als der Affe und dieser wiederrum als der Mensch? Hier geht es nicht darum, den Menschen „nur“ als irgendein Vieh abzustempeln, der über keine besonderen Merkmale verfügt, die ihn von allen anderen Lebewesen unterscheiden. Es geht um rein evolutionäre Fragestellungen: Der Erfolg der Evolution misst sich danach sich an die Umwelt anzupassen und seine Gene an die Nachkommen weiterzugeben. Weil heute Regenwurm, Schimpanse und Mensch existieren, haben sie sich auf ihre Weise unterschiedlich an die Umwelt angepasst und sich fortgepflanzt. Man könnte zwar einwenden, dass der Schimpanse klüger ist als der Regenwurm, gleichzeitig kann man aber mit Fug und Recht behaupten, dass Regenwürmer in ihrer Fortpflanzung und Anpassung an die Umwelt viel erfolgreicher sind. Weil alle heute lebenden Arten bis in unsere Gegenwart existieren, kann man daher davon ausgehen, dass sie ihre eigenen evolutionären Strategien hatten, die sie erfolgreich meistern konnten.
Es gibt also keine höheren oder niedrigen entwickelten Arten. Da wir alle auf einen gemeinsamen Vorfahren zurückgehen, haben alle heutigen Organismen die gleiche Evolutionsgeschichte hinter uns. Man hat sich nur in unterschiedliche Richtungen hin entwickelt und spezialisiert.
Evolutionslinien verlaufen nicht in einer Linie, sondern sie verzweigen sich an vielen Stellen, sodass mehrere Entwicklungslinien parallel verlaufen. Das ist Folge des Evolutionsprozesses: einzelne Populationen spalten sich, z. B. in Form von geographischer Isolation, von der Gründerpopulation ab. So entstehen aus einer Population zwei oder mehr. Mit ausreichend Zeit und Isolation, also dass sich die Populationen nicht mehr vermischen können, entwickeln sie sich unabhängig voneinander weiter zu neuen Arten. Wiederum können sich aus diesen Populationen durch Isolation weitere abspalten und neue Arten bilden. Durch diese Artabspaltung verläuft die Evolution also nicht auf einer Linie, oder Leiter, sondern als Busch mit vielen Abzweigungen, Seitenlinien, Parallelentwicklungen und Aussterbeereignissen (Abb. 7).
Abb. 7: die Evolution ist keine Leiter von niederen zu höheren Lebewesen, sondern verläuft „buschartig“.
Es treiben ständig neue Äste hervor, alte sterben ab. Doch kein Zweig bevorzugt eine bestimmte Richtung. Arten können sich aufspalten und dann in verschiedene Richtungen weiterentwickeln.
Tatsache ist also: evolutionäre Übergänge im Fossilbericht sind gut dokumentiert. Ich habe mich hier auf einige wenige populäre Fossilien wie Archaeopteryx und Tiktaalik konzentiert – doch es gibt eine ganze Reihe verschiedenster Formen. Entscheidende evolutionäre Übergänge im Fossilbericht werden in den kommenden Episoden vorgestellt.
Literatur
AG EVOLUTIONSBIOLOGIE (NEUKAMM, M.): Die evolutionäre Zwischenform Tiktaalik roseae https://www.ag-evolutionsbiologie.net/html/2008/tiktaalik.html
AG EVOLUTIONSBIOLOGIE (HEMMINGER, H.): Doch noch in Georgien angekommen: Die Fossilien von Dmanissi aus Sicht des Kreationismus https://www.ag-evolutionsbiologie.net/html/2015/wie-wort-und-wissen-uebergangsformen-wegdefiniert.html
AHLBERG, P. E. & CLACK, J. A. (2006) A firm step from water to land. Nature 440, S. 747-749.
DAESCHLER, E. B.; SHUBIN, N. H. & JENKINS, F. A. (2006) A Devonian tetrapod-like fish and the evolution of the tetrapod body plan. Nature 440, S. 757-763.
DALTON, R. (2006) The fish that crawled out of the water. A newly found fossil links fish to land-lubbers. www.nature.com/news/2006/060403/full/060403-7.html.
FUTUYMA, D. J. (1990) Entwicklungsintegration und Makroevolution. In: ders.: Evolutionsbiologie. Birkhäuser, Basel, S. 497-498.
GOULD, S. J. (2002) Illusion Fortschritt. Die vielfältigen Wege der Evolution. Fischer, Frankfurt.
JUNKER, R. (2006) Tiktaalik – ein erstklassiges Bindeglied? www.genesisnet.info/index.php?News=63
JUNKER, R. & SCHERER, S. (1998) Evolution. Ein kritisches Lehrbuch. Weyel, Gießen.
KUTSCHERA, U. (2006) Evolutionsbiologie (2. erweiterte Auflage). Eugen Ulmer, Stuttgart.
MAHNER, M. (1986) Kreationismus – Inhalt und Struktur antievolutionistischer Argumentation. Pädagogisches Zentrum, Berlin.
MAYR, E. (1967) Artbegriff und Evolution. Berlin
MILLER, K. B. n.d. Taxonomy, transitional forms, and the fossil record. http://www.asa3.org/ASA/resources/Miller.html
NEDIN, C. (1999): All about Archaeopteryx http://www.talkorigins.org/faqs/archaeopteryx/info.html
NEUKAMM, M. & KUTSCHERA, U. (2006) Zwischenformen und Modellsysteme der Evolutionsbiologie. www.martin-neukamm.de/mudskipper.pdf
PROTHERO, D. (2017): Evolution – What the Fossils say and why it matters. Second edition. New York: Columbia University Press
SHUBIN, N. H.; DAESCHLER, E. B. & JENKINS, F. A. (2006) The pectoral fin of Tiktaalik roseae and the origin of the tetrapod limb. Nature 440, S. 764-771.
TALKORIGINS: Archaeopteryx http://www.talkorigins.org/indexcc/CC/CC214_1_1.html
