

Kreationisten und das Grundtypenmodell
ein Großteil des Textes basiert auf Martin Neukamm: Die kreationistische Grundtypenbiologie in der Kritik
Unser blauer Planet ist voller Leben. Heute sind etwa 2 Mio. Tier- und Pflanzenarten wissenschaftlich beschrieben worden.
Wir wissen, dass neue Arten durch Evolution immer wieder aufs Neue entstehen oder aussterben.
Religiöse Fundamentalisten sind da aber anderer Ansicht.
Text als pdf
Sie beharren darauf, dass die Vielfalt des Lebens nicht durch Evolution erklärt werden kann. Stattdessen seien alle Arten durch einen göttlichen Schöpfungsakt entstanden. Aber bei der Menge an beschriebenen Arten stehen sie vor einem kleinen Dilemma biblischen Ausmaßes. Weil Gottes höchste Schöpfung nun mal unerträgliche Sünder waren, kam es zur Sintflut. Aber Gott wählte einen ganz besonderen Mann aus – Noah – der sich und seine Familie retten soll. Dafür soll er ein großes Schiff bauen, die Arche, und von jedem Tier jeweils ein Paar auf diese Arche bringen. Von einigen Tieren sollen es sogar sieben Paar sein dürfen. Auch wenn die Arche mit 155m Länge sicherlich kein kleines Schiff war, wäre es bei so vielen Tierarten ganz schön eng geworden. Das haben selbst die leichtgläubigsten Kreationisten gemerkt.
Laut einiger Kreationisten ist es gar nicht notwendig gewesen, alle Arten auf die Arche zu bringen, denn Gott habe nicht alle Arten, sondern gewisse „Grundtypen“ erschaffen, im englischen auch „kinds“ genannt. Gebräuchlich ist auch der Begriff der Baraminologie. Gemäß des Grundtypmodells sollen alle Lebewesen durch Mikroevolution aus solchen Grundtypen hervorgegangen sein, die im Kreationismus als das Ergebnis göttlicher Schöpfung gedeutet werden. Der Schöpfer habe gewisse Grundtypen erschaffen, die bereits den kompletten Merkmalsbestand der heute existierenden Artenwelt bzw. das genetische Potential zu dessen Entstehung enthielten. Hinzu wird angenommen, dass aus einem Grundtyp niemals ein anderer entstehen könne. Die Grundtypen haben keine evolutionäre Verbindung und können sich untereinander nicht fortpflanzen.
So gibt es den Grundtyp „Hund“, „Katze“, „Affe“, „Mensch“ usw.
Aus dem Grundtyp „Hund“, quasi gesehen als Stammart, haben sich dann alle Hundeartigen entwickelt: Wolf, Streifenschakal, Rotfuchs etc. Eine sog. Evolution innerhalb dieser Grundtypen sei also möglich, jedoch nicht darüber hinaus (Abb. 1).
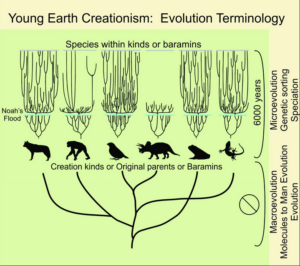
Abb. 1: Grundtypenmodell des Kreationismus
Diesen so genannten „genetisch polyvalenten“ Stammarten wird demnach ein eingeschränktes, „programmiertes“ Variationspotential zugebilligt: Sie sollen sich im Laufe der Zeit durch „Mikroevolution“ – d.h. durch „Variationsvorgänge auf der Basis bereits vorhandener Konstruktionen“ (Junker 2002, S. 14) – zu den gegenwärtigen Formen, Rassen oder Arten ausdifferenziert haben, die jeweils einer gemeinsamen Schöpfungseinheit angehören. Indem die Merkmale variieren und mehr oder minder unsystematisch auf die Nachkommen verteilt werden, soll also eine gewisse Artenvielfalt entstehen. Der Erwerb qualitativ neuer Merkmale (Makroevolution), welcher der Schaffung eines neuen Grundtyps analog wäre, wird hingegen als unplausibel erachtet, zumal angenommen wird, dass diese Aufgabe allein dem Schöpfer zufalle. Damit stünden die Arten verschiedener Grundtypen in keiner Relation der gemeinsamen Abstammung, und infolge fortschreitender Artbildung und Spezialisierung würde das genetische Variationspotential in der Tendenz langsam abnehmen.
Das Grundtypmodell wird nun von den Kreationisten gerne als Aushängeschild eines prüfbaren und damit wissenschaftlichen Schöpfungsmodells angeführt.
Ist aber dieses kreationistische Grundtypenmodell aber eine alternative zur Evolutionstheorie?
Wissenschaftsphilosophische Überlegungen
Wissenschaftliche Theorien können nicht ohne weiteres direkt getestet werden, denn sie beziehen sich in der Regel auf nicht direkt beobachtbare Sachverhalte und sagen zudem nichts Spezielles aus, sondern beschreiben bzw. erklären ihren Gegenstandsbereich nur allgemein.
Folglich müssen die allgemeinen Aussagen spezifiziert werden, d.h., man muss aus der allgemeinen Theorie ein mehr oder weniger spezifisches Modell gewinnen, um zu Aussagen über beobachtbare Sachverhalte zu gelangen. Dazu muss die Theorie mit Zusatzannahmen (Hilfs- und Indikatorhypothesen) angereichert werden, wobei diese zunächst die Eigenschaften der von der Theorie behandelten Objekte konkretisieren und den Zusammenhang mit beobachtbaren Sachverhalten herstellen (Mahner & Bunge 2000).
Betrachten wir beispielsweise die Evolutionstheorie, stellen wir fest, dass sie nur Aussagen über die Evolution der Lebewesen allgemein enthält, nicht jedoch ohne Weiteres Aussagen über die Entstehung einer bestimmten Spezies. Will man hier im Rahmen eines Modells zu prüfbaren Aussagen über die Evolution einer konkreten Art gelangen, muss das allgemeine Erklärungsschema etwa mit zusätzlichem Wissen über die strukturellen, funktionellen oder entwicklungsbiologischen Details der betreffenden Spezies versehen und eventuell mit selektionstheoretischen sowie historischen Zusatzannahmen ergänzt werden.
Analog dazu lässt sich auch aus dem Darwin’schen Postulat, wonach alle Arten von einem gemeinsamen Vorfahren abstammen, zunächst noch nichts Konkretes folgern. Erst in Verbindung mit zusätzlichem Wissen über die Mechanismen der Vererbung, Variation und Selektion lässt sich die konkrete Folgerung ableiten, dass dann zwischen den Arten eine abgestufte Ähnlichkeit bestehen muss; eine Folgerung, die man durch Beobachtung testen kann und die bis zur molekularen Ebene immer wieder bestätigt wird.
Entscheidend ist nun für unsere Argumentation, dass die Zusatzannahmen in den Naturwissenschaften nicht etwa willkürlich gewählt werden dürfen. Vielmehr muss jede Hilfshypothese, mit der eine Theorie versehen wird, unabhängig davon prüfbar, empirisch wohlbegründet sowie mit unserem Hintergrundwissen kompatibel sein (Kitcher 1982; Mahner & Bunge 2000). Dies ist bei den oben genannten Beispielen auch der Fall: Alle zusätzlichen Vorstellungen bezüglich der Vererbung, Variation und Selektion sowie über die Struktur und Entwicklung bestimmter Organismen lassen sich im Rahmen der unterschiedlichen biologischen Disziplinen testen und belegen, ohne dass hierfür die Evolutionstheorie vonnöten wäre.
Wie aber sieht es unter diesem Gesichtspunkt mit dem Kreationismus und seinem Grundtypmodell aus? Auch hier gilt, dass die allgemeine Schöpfungshypothese zunächst mit Zusatzannahmen angereichert werden muss, damit sich aus ihr prüfbare Folgerungen ergeben. Im Rahmen entsprechender Hilfshypothesen wird dann eben behauptet, der Schöpfer habe genetisch polyvalente Stammarten erschaffen, sie gemäß eines „Baukastenprinzips“ einander ähnlich gemacht und Mikroevolution innerhalb abgrenzbarer Grundtypen zugelassen.
Entgegen der Auffassung ihrer kreationistischen Protagonisten ist es allerdings nicht möglich, eine Schöpfungs-Forschung zu betreiben bzw. die Schöpfungshypothese durch den erfolgreichen Test der theoretischen Erwartungen plausibel zu machen. Weshalb nicht? Weil die hierzu erforderlichen Zusatzannahmen allesamt beliebig und darüber hinaus gar nicht unabhängig von der Schöpfungshypothese prüfbar sind! So ist beispielsweise die Annahme, dass die Lebewesen mit einer „programmierten“, aber schlussendlich begrenzten Variabilität ausgestattet wurden, keineswegs zwingend; auch der gegenteilige Befund der nahezu unbegrenzten Variabilität und makroevolutionären Entwicklung der Arten könnte Teil des Schöpferplans sein. Immerhin nehmen ja nicht wenige religiöse Menschen an, die Evolution sei die „Methode der Schöpfung“, kommen also zu einem ganz entgegengesetzten, ebenso willkürlichen Schluss.
Ebenso lässt sich die Frage, weshalb der Schöpfer seine Arten ausgerechnet nach einem Baukastenprinzip erschaffen hat, anstatt jede Art bis ins kleinste Detail völlig verschieden zu gestalten, auf empirischem Wege nicht beantworten. Könnte man nicht genauso gut annehmen, dem Schöpfer habe es gefallen, ein Formenkontinuum hervorzubringen? Bei dieser Annahme würde sich allerdings die Schöpfungsfrage im Rahmen des Grundtypmodells gar nicht mehr stellen. Man sieht hier also, dass die kreationistischen Hilfshypothesen willkürlich so gewählt wurden, um ein bestimmtes Schöpferbild empirisch plausibel erscheinen zu lassen, das es ohne diese Annahmen gar nicht wäre. Man kann in diesem Punkt noch weiter gehen und fragen, ob sich ein übernatürliches Wesen überhaupt an weltimmanente Regeln halten und sich den aus den Naturgesetzen resultierenden Zwängen unterwerfen muss. Hätte ein Schöpfer, dem es gefiel, Wasser in Wein zu verwandeln, nicht auch Arten erschaffen können, die auf der Sonne leben, ohne Nahrung auskommen oder aus Luft bestehen, anstatt das Gesetzesnetz der Natur so und nicht anders vorherzubestimmen, wie wir es kennen (Neukamm 2004)?
Alles in allem zeigt sich, dass sich für das Wirken übernatürlicher Wesen keine objektive Grenze angeben lässt.
Verzichtet man auf solche Hilfshypothesen und akzeptiert nur Zusatzannahmen, die sich unabhängig von der Schöpfungshypothese testen lassen und die empirisch wohlbestätigt sind, lässt sich die Schöpfungshypothese nicht mehr evident machen. Egal, welche Forschungsergebnisse man auch immer zu Tage fördert, die Schöpfungshypothese wäre von einer empirischen Bestätigung des Grundtypmodells nicht betroffen, da sich über die Schöpfungsakte übernatürlicher Wesen nach Belieben spekulieren lässt. Die Schöpfungsthese steckt gewissermaßen in der logischen Isolation, ist ohne willkürlich getroffene Zusatzannahmen mit allen nur denkbaren Beobachtungen kompatibel und somit grundsätzlich unfalsifizierbar.
Erschwerend kommt hinzu, dass sich die Entwicklung der Arten, wie sie das Grundtypmodell beschreibt, nur vor einem strikt naturalistischen Hintergrund erforschen und beschreiben lässt, d.h., wir müssten auf natürliche Mechanismen und Gesetzmäßigkeiten Bezug nehmen, um die im Grundtypmodell beschriebenen Vorgänge sowie die evolutionären Beschränkungen zu verstehen.
So kann das Grundtypmodell aufgrund der Annahme, ein Schöpfer habe genetisch isolierte Stammformen erschaffen, eine Reihe von Fragen nicht mehr beantworten, die durch Darwins Theorie ganz elegant beantwortet werden (Korthof 2004). Warum existieren z.B. zwischen den Arten verschiedener Grundtypen bemerkenswerte Ähnlichkeiten, wenn zwischen ihnen kein stammesgeschichtlicher Zusammenhang besteht? Die Grundtypen „Hunde“ und „Katzen“ lassen sich anhand bestimmter Merkmale in die Ordnung der „Carnivora“ eingliedern, welche dann zusammen mit anderen Grundtypen wie „Pferde“ zu Säugetieren gruppieren lassen und diese zusammen mit anderen Grundtypen, z. B. „Entenvögel“ zu Wirbeltieren gruppieren usw.
Weshalb bietet die Annahme der gemeinsamen Abstammung der Arten innerhalb von „Grundtypen“ eine gute Erklärung, für höhere Kategorien hingegen nicht? Wenn Mensch und Affe verschiedene Grundtypen seien, wie kann es sein dass der „Affe“ „Schimpanse“ dem Grundtypen „Mensch“ genetisch näher steht, als dem „Gorilla“, obwohl dieser , wie der Schimpanse, zum Grundtyp „Affe“ gehört?
Mit anderen Worten: Wie lässt sich die Formen-Hierarchie der Organismen-Gruppen überhaupt noch konsistent erklären?
Wer an dieser Stelle darauf verweist, der Schöpfer habe eben nach einem „Baukastenprinzip“ Grundtypen mit „mehr oder weniger zahlreichen Gemeinsamkeiten“ erschaffen, tut dies wie oben betont aufgrund willkürlicher Zusatzannahmen, die nur dazu dienen, ein bestimmtes Schöpfer- bzw. Gottesbild zu retten. Im Gegensatz dazu lässt die Evolutionstheorie auf der Grundlage der Mechanismen Variation, Selektion und Vererbung, die hier als legitime (weil empirisch gesicherte) Zusatzannahmen mit der Deszendenzhypothese verknüpft werden, gar keinen anderen Befund erwarten und liefert daher im Gegensatz zum Schöpfungsmodell auch eine spezifische Erklärung für die abgestuften Ähnlichkeiten jenseits der „Grundtypen“.
Ein weiteres Erklärungsproblem verbindet sich mit der Frage, wie etwa die enorme genetische Vielfalt der 65 000 Arten des Grundtyps Rüsselkäfer oder alle Merkmale der Arten des Grundtyps der Fasanenartigen in einer einzigen Stammart untergebracht worden sein sollen (Korthof 2004).
Wenn Kreationisten behaupten, dass zahlreiche Merkmale der Arten eines Grundtyps, wie z.B. „Schillerfarben, Krönchen, Pfauenauge und dergleichen“, in der Stammform des „Fasanenartigen-Grundtyps“ noch nicht „phänotypisch ausgeprägt“ waren, ist fraglich, ob man sie überhaupt „zum ursprünglichen Bestand der polyvalenten Stammform“ rechnen kann (Junker 2005). Denn wenn sich derartige Merkmale erst sukzessive durch evolutionäre (!) Mechanismen wie Mutation, Selektion, Genrekombination usw. herausgebildet haben, hätte der Schöpfer streng genommen nicht sie, sondern nur die genetische Prädisposition bzw. das Potenzial zu ihrer Entstehung erschaffen.
Die erstaunlichsten und unterschiedlichsten Ornamente im Tierreich entstehen als Folge von Mutation und Selektion – Ornamente, die man sicher mit Recht als qualitative Neuheiten einstufen darf. Das aber wäre ja schon jene Makroevolution der Evolutionsbiologie, die im Rahmen des Grundtypmodells als Mikroevolution deklariert wird, um nicht zugeben zu müssen, dass evolutionäre Neuheiten nicht anders als durch Variation bereits vorhandener Strukturen entstehen. Überhaupt ist das Ausschöpfen der im epigenetischen System angelegten Entwicklungspotenzen bzw. das Neuarrangement konservierter Merkmale und Regulationsmechanismen zunehmend als Ursache makroevolutionärer Veränderungen in der Diskussion (s. z.B. Lorenzen 1988, S. 932; Hall 1996, S. 227-229; Kirschner & Gerhart 2005); es bedarf hierzu nicht zwingend der Erfindung neuer Gene (Ridley 2003).
Wir wissen z. B., dass viele Gene, die die Embryonalentwicklung der Tiere steuern, hoch konserviert sind. Genannt seien hier die Hox-Gene, die in allen mehrzelligen Tieren vorkommen. Primäre Aufgabe der Hox-Gene ist die Gliederung des Embryos entlang der Körperlängsachse. Sie liefern den Zellen des Embryos Lageinformationen und Informationen über ihre weitere Teilung, ihre Differenzierung und ggf. ihr programmierter Zelltod (Apoptose). Hox-Gene sind inzwischen bei allen Bilateria nachgewiesen worden, so auch bei Insekten und Wirbeltieren (vgl. Carroll 2008; Abb. 2).
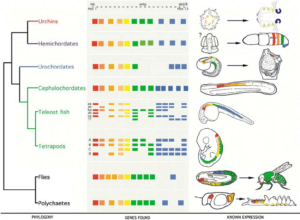
Abb. 2: Hox Gene
Die Hox-Gene der verschiedenen Tiergruppen unterscheiden sich selbst nur an wenigen Stellen. Ihre Genexpression, also zu welchem Zeitpunkt und an welcher Stelle die Gene an- oder ausgeschaltet werden, ist aber unterschiedlich. Während der Evolution sowohl von Gliederfüßern als auch von Wirbeltieren korreliert die Veränderung der Expression von Hox-Genen mit der Veränderung der Körperbaupläne (Anlageplan).
Hier haben wir also, wenn wir es so sagen wollen, dasselbe genetische Potential, um komplexe Körperformen zu erzeugen. Würde dies wiederum nicht bedeuten, dass man eigentlich nur einen Grundtyp der Tiere braucht? Den gemeinsamen Vorfahren aller Tiere – und wir wären wieder bei der Evolution.
Wie neu muss also eine qualitative Neuheit sein, damit sie auch Evolutionsgegner als „echte“ Neuheit (Junker 2005) anerkennen? Wird die Unterscheidung zwischen Mikro- und Makroevolution im Rahmen des Grundtypmodells überhaupt konsistent getroffen, oder werden die Begriffe so verwendet, um das Auftreten evolutionärer Neubildungen von vornherein wegzudefinieren?
Damit nicht genug: natürlich treten in der Evolution weitere Mutationen auf, die in der Stammart nicht vorhanden sind.
Die Züchtungen der Art Brassica oleracea, dem Gemüsekohl, führten zu verschiedensten Gemüsesorten: Kohl, Brokkoli, Blumenkohl und Kohlrabi (Abb. 3). Hier führten graduelle Akkumulationen kleiner Mutationen zu völlig verschiedenen Gemüsesorten. Sie zählen aber zu einer Art und zum selben Pangenom. Das Pangenom bezeichnet die Gesamtheit der Gene in einer Art. Das Pangenom umfasst zwei Untergruppen: Das Kerngenom, das die Gene umfasst, die in jedem Mitglied der Spezies vorkommen und das variable (akzessorische) Genom, das die Gene beinhaltet, die nur in einzelnen Mitgliedern der Art vorhanden sind (Brockhurst et al. 2019). All diese Züchtungen entstammen also demselben genetischen Ursprung, sie sind lediglich eine Modifikation dieses Pangenoms. Diese Gemüsesorten entstanden durch künstliche Selektion durch den Menschen, indem sie nach Blattform, Blütenfarbe usw. selektierten. Diese Variationen waren das Ergebnis von Mutationen, die die neuen Variationen haben und so von ihren Ursprungsvariationen zu unterscheiden sind. Das ist Evolution: Jede neu gebildete Gruppe unterscheidet sich durch für sie einzigartige Merkmale, die sich auch genetisch zeigen. Und das führt beim kreationistischen Grundtypenmodell zu einem Problem: Nach diesem müssten in den Grundtypen nämlich alle potentiellen Variationen im Genom schon vorhanden sein. Entsprechend müsste bei Brassica oleracea das gesamte genetische Programm für all seine Sorten vorhanden sein. Aber das trifft eben nicht zu! Genomische Analysen des Gemüsekohls und seiner Zuchtformen zeigen, dass 20% der Gene in diesem Pangenom nur in einigen der Untergruppen vorkommen. Diese Gene hängen mit wichtigen agronomischen Merkmalen wie Blütezeit und Krankheitsresistenzen zusammen (Golicz et al. 2016).
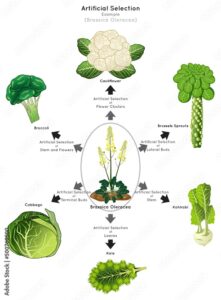
Abb. 3: Brassica oleracea
Ähnliches auch bei Hunden und Wölfen: Man findet zwar gemeinsame genetische Merkmale zwischen Wölfen und Haushund, aber man kann wichtige genetische Merkmale eines Dackels oder Pitbulls nicht im Wolf finden! Weil die Haushundrassen vom Wolf abstammen und neue genetische Variationen entwickelten, die im Wolfsgenom nicht vorhanden sind (Lindblad-Toh et al. 2005).
Wer hier auf die Idee kommt, das in den Genen schlummernde Entwicklungspotenzial in den Plan des Schöpfers einzubeziehen, der in weiser Voraussicht gezielt „vorteilhafte Entwicklungsrouten“ angelegt habe, sieht sich im Übrigen mit absurden Konsequenzen konfrontiert. Er müsste nach Darwin ja dann auch annehmen, „dass Gott die unzähligen Variationen der Haustiere und Kulturpflanzen speziell für den Nutzen der Züchter vorherbestimmt habe; dass beispielsweise Kropf und Schwanzfedern der Tauben variieren, damit die Taubenliebhaber ihre grotesken Formen züchten können, und dass Hunde in ihren geistigen Fähigkeiten variieren, damit man Kampfhunde züchten könne“ (Junker 2004, S. 11). Diese Folgerungen dürften auch Kreationisten abwegig erscheinen. Sie sind jedoch nicht weniger kurios als die Annahmen, mit denen die Kreationisten ihr Schöpfungsmodell ausstatten.
Die wissenschaftliche Irrelevanz des Grundtypmodells
Unabhängig von den methodologischen Problemen der Schöpfungshypothese wird oft behauptet, das Grundtypmodell könne eine Reihe von Befunden erklären, die im Rahmen der Evolutionstheorie problematisch oder zumindest wenig plausibel erscheinen (Junker 2005). Tatsächlich aber werden die meisten Befunde gar nicht exklusiv durch das Grundtypmodell erklärt; es handelt sich in den meisten Fällen um Erwartungen moderner entwicklungsbiologischer Konzepte und Evolutionstheorien. Wie wir gesehen haben, ist z.B. eine „programmierte Variabilität“ in gewissem Umfang auch im Rahmen der Evolutionsbiologie zu erwarten, und die durch Kreuzungsexperimente feststellbare „Abgrenzbarkeit“ der Arten auf der Ebene bestimmter systematischer Einheiten („Grundtypen“) deutet einfach nur darauf hin, dass sich die Spezies über einen langen Zeitraum evolutionär auseinanderentwickelt haben, sodass keine Kreuzungen mehr möglich sind.
Auch die Tatsache, dass Organismen nur eingeschränkt variabel sind, ist für die Evolutionsbiologie wenig überraschend, weil angesichts funktioneller Bürden und konstruktiver Zwänge immer nur ganz bestimmte Formen tauglich sind und die Entwicklung streckenweise in vorgegebene Bahnen gelenkt wird (s. Futuyma 1990, S. 497-498; Riedl & Krall 1994, S. 263). Dieses Prinzip der „funktionellen Bürde“ (vgl. Riedl 1990, 218ff; Neukamm 2014, 105) ist inzwischen gut erforscht und kann an vielen Beispielen nachvollzogen werden.
- Von der Effizienz des vierkammerigen Herzens der Säugetiere hängt die Funktion so vieler Merkmale und Organe ab, dass sich seine Struktur nicht mehr nennenswert verändern kann.
- Wale und Delfine rekapitulieren während ihrer Entwicklung den konventionellen Säugerbauplan mit Zahnanlagen und Beinknospen, obwohl sie weder Kauwerkzeuge noch Extremitäten besitzen. Der Grund ist, dass die Organanlagen für eine formgerechte Entwicklung des Kiefers und des Beckens benötigt werden.
- Bei den Wirbeltieren bilden sich während der Embryonalentwicklung aus dem sogenannten Neuralrohr Gehirn, Rückenmark und Augen. Unter der „organisierten Herrschaft“ des „Mastergens“ PAX-6 quellen aus dem hinteren Bereich des Vorderhirns Wülste (die sog. Augenblasen) hervor, die sich zum doppelwandigen Augenbecher umformen. Die Ausstülpung, aus der die künftige Netzhaut entsteht, kann nur in „eine Richtung“ erfolgen. Aufgrund dieses konstruktiven Zwangs wird die Netzhaut ebenfalls der Inversion unterzogen, obwohl dies konstruktiv widersinnig ist.
Wir sehen also, dass die „kausalen Mechanismen für die Invarianz der Entwicklung“ entwicklungsbiologische Mechanismen sind, durch die sich die Abgrenzbarkeit der Baupläne begründet. Aus Sicht der Evolution, die ihr stammesgeschichtliches Erbe nicht abschütteln kann, ist die starre Rekapitulation des Säugerbauplans bei Walen, die inverse Netzhaut bei Wirbeltieren u.v.a. zwingend. Ihre Entstehung wurde durch historisch gewachsene Entwicklungsprozesse, die sich teils bis zu den urtümlichen Chorda-Tieren zurückverfolgen lassen, erzwungen.
Infolgedessen kommt es in der Evolution zu keinem stetigen, harmonischen Wandel aller Merkmale, sondern zu unabhängiger Entstehung bauähnlicher Merkmale (Konvergenzen) sowie zu mosaikartigen Diskontinuitäten im Merkmalsgefüge der Organismen. Es ist demnach keineswegs so, dass evolutionstheoretisch gesehen eher ein Formenkontinuum zwischen eng verwandten Arten zu erwarten sei, während umgekehrt die systematisch auftretenden „Diskontinuitäten zwischen benachbarten Grundtypen“ eine Schwächung der Evolutionstheorie zu Gunsten der Schöpfungsthese bedeuteten (Junker 2005). Die Evolution hat eben Grenzen, die epigenetisch zu verstehen sind und keineswegs auf die Existenz eines transnaturalen Zwecksetzers hindeuten.
Wie viele Grundtypen waren es denn nun?
Versuchen wir aber ernst zu bleiben und fragen: Wie viele Grundtypen brachte denn nun Noah auf die Arche?
Laut Ken Ham, dem Gründer der Arc Encounter, also dem fundamentalistischen Freizeitpark für Kreationisten, welches eine lebensgroße Arche Noah in Kentucky zeigt, lebten in der Arche 6.700 Tiere, die 1.400 Grundtypen darstellen (von manchen Grundtypen gab es sieben Paare statt eines).
In Wirklichkeit schafft diese Minimierung der Anzahl der Tiere auf der Arche ganz neue Probleme.
Also: aus 1.400 Grundtypen sind die heute etwa 2 Mio. beschriebenen Arten entstanden – und zwar in einem Zeitraum von 4.300 Jahren! Das wäre eine super-mega-ultraschnell-Evolution! Nun gut, Ken Ham wendet ein, dass ja die Wassertiere nicht mitgenommen wurden und wahrscheinlich auch nicht die Insekten. Pflanzen sowieso nicht. Hier besteht aber das Problem, dass Süßwasserorganismen sterben, wenn sie im Salzwasser sind und anders rum. Wenn Insekten nicht in die Arche mitgenommen wurden, können wir davon ausgehen, dass die meisten von ihnen dann ausgestorben sind – sie sind entweder ertrunken oder verhungert – und wurden danach wohl neu erschaffen? Auch hier die Frage: jedes der 1 Mio. bisher bekannten Insektenarten einzeln? Oder doch nur einen Grundtypen Insekt? Schließlich ist jede Insektenart eine abgewandelte Form vorheriger Insekten. Und alle Insekten sind abgewandelte Arthropoden und alle Arthropoden sind abgewandelte Ecdysozoa und jedes Ecdysozoa ist ein abgewandelter Bilateria. Warum also nicht gleich einfach einen ursprünglichen Einzeller nehmen? Es gibt entgegen kreationistischen Behauptungen keine kategorialen Unterschiede zwischen höheren Kladen. Dasselbe gilt auch für Pflanzen und Pilze. Die wenigsten Arten hätten die Flut überlebt. Aber gut, gehen wir davon aus, dass in der Arche nur Landwirbeltiere genommen wurden. Von denen gibt es immerhin etwa 35.000 heute lebende Arten: innerhalb von 4.000 Jahren von 1.400 Grundtypen auf 35.000 Arten zu kommen ist ein starkes Stück und übersteigt jede Erwartungshaltung der Evolution. Sind Kreationisten nun Hyper-Evolutionisten? Außerdem: laut Ken Ham wurden ja jeweils sieben Paare von bestimmten Haustieren genommen. Aber warum sollten z. B. Rinder und Schafe zwei verschiedene Grundtypen sein? Beide gehören zu den sog. Hornträgern, zusammen mit Büffeln, Ziegen und Antilopen.
Nicht zu vergessen: Laut Ken Ham gab es auch Dinosaurier auf der Arche, kann man sogar im Arc Encounter bestaunen. Wir müssen also die ausgestorbenen Landwirbeltiergruppen mit einbeziehen, von denen mindestens genauso viele beschrieben wurden, wie heute existieren. Vom Sinn und Unsinn, unzählige Arten (oder Grundtypen?) mitzuführen, die kurz darauf aussterben werden, mal ganz zu schweigen… Es gibt natürlich noch viele weitere Probleme, wie die geographische Verbreitung der Arten und die Tatsache, dass Noah weder Amerika noch Australien und die dortige Tierwelt kannte – aber gut, belassen wir es dabei.
Wir sehen: Das ganze Thema „Baraminologie“ erinnert an eine lächerlich schlechte Nachahmung der Wissenschaft – Wissenschaft, wie sie sich Kinder beim Spielen vorstellen, wo sie wissenschaftlich klingende Worte sagen, die überhaupt keinen Sinn ergeben. Der Fokus kreationistischer „Forschung“ besteht darin, das gesamte Gebiet der modernen Taxonomie zu umgehen und sich dann Möglichkeiten auszudenken, Hunderte von einzelnen Arten und Gattungen in möglichst wenige Kategorien einzuteilen. Sie machen sich nicht die Mühe, mit echten Tieren zu arbeiten, machen sich nicht die Hände schmutzig mit anatomischen Arbeiten, die die moderne Taxonomie begründet haben und haben nicht den Willen, jahrelang in den Universtäten zu arbeiten, um die Art von Ausbildung zu erhalten, die zum Verständnis und zur Analyse der molekularen Phylogenetik erforderlich ist und geben sich nicht die Mühe die gigantische Literatur der modernen Taxonomie und Kladistik zu durchforsten. Nein, das würde erfordern, dass sie in der wissenschaftlichen Arbeit geschult werden und sich der Tatsache der Evolution stellen müssen, die sich durch die ganze Lebenswelt zieht. Stattdessen führen sie sehr oberflächliche Analysen auf niedrigem Niveau durch, bei denen sie hier und da Ideen aus stark vereinfachten Internetquellen herauspicken.
Literatur
Brockhurst, M. A. et al. (2019): The Ecology and Evolution of Pangenomes. In: Curr Biol. Band 29, Nr. 20, 2019, S. R1094–R1103. doi:10.1016/j.cub.2019.08.012
Carroll, S. B. (2008): Evo Devo. Das neue Bild der Evolution. Berlin University Press
Futuyma, D.J. (1990): Entwicklungsintegration und Makroevolution. In: ders.: Evolutionsbiologie. Birkhäuser, Basel, 497-498.
Golicz, A., Bayer, P., Barker, G. et al. The pangenome of an agronomically important crop plant Brassica oleracea. Nat Commun 7, 13390 (2016). https://doi.org/10.1038/ncomms13390
Hall, B.K. (1996): Baupläne, Phylotypic Stages, and Constraint. Why There are so few Types of Animals. Evolutionary Biology 29, 215-261.
Junker, R. (2002): Ähnlichkeiten, Rudimente, Atavismen. Design-Fehler oder Design-Signale? Hänssler, Holzgerlingen.
Junker, R. (2005): Kritik an der Grundtypenbiologie. www.genesisnet.info/index.php?Artikel=1246&Sprache=de&l=2
Junker, T. (2004): Evolution und die Marmeladetheorie des Erdkerns. Materialien und Informationen zur Zeit 33(3), 10-13.
Kirschner, M.W.; Gerhart, J.C. (2005): The Plausibility of Live Resolving Darwin’s Dilemma. Yale University Press: New Haven, CT.
Kitcher, P. (1982): Abusing Science: The Case Against Creationism. MIT-Press, Cambridge, MA.
Korthof, G. (2004): Common Descent. It’s All or Nothing. In: Young, M.; Edis, T. (Hrsg.): Why Intelligent Design Fails. A Scientific Critique of New Creationism. Rutgers University Press: New Brunswick, NJ, 32-47.
Lindblad-Toh, K., Wade, C., Mikkelsen, T. et al. Genome sequence, comparative analysis and haplotype structure of the domestic dog. Nature 438, 803–819 (2005). https://doi.org/10.1038/nature04338
Lorenzen, S. (1988): Die Bedeutung synergetischer Modelle für das Verständnis der Makroevolution. Eclogae Geol. Helv 81(3), 927-933.
Mahner, M.; Bunge, M. (2000): Philosophische Grundlagen der Biologie. Springer: Berlin.
Neukamm, M. (2004): Kreationismus und Intelligent Design. Über die wissenschaftsphilosophischen Probleme von Schöpfungstheorien. www.martin-neukamm.de/kreation.pdf
Neukamm, M. (2005) Die kreationistische Grundtypenbiologie in der Kritik. Warum es keine empirisch-wissenschaftliche Schöpfungsforschung geben kann. Skeptiker 18 (4/05), 144–150.
Neukamm, M. (2014) Darwin heute: Evolution als Leitbild in den modernen Wissenschaften. WBG, Darmstadt.
Neukamm, M. (ohne Jahr): Schöpfung: Die Grundtypen des Kreationismus. Kritik unerwünscht. Erläuterungen zum Grundtypmodell https://www.ag-evolutionsbiologie.net/html/2015/grundtyp-modell-verfehlte-kritik.html
Ridley, M. (2003): What Makes You Who You Are: Which Is Stronger – Nature or Nurture? Time 161(22), 54-63.
Riedl, R. (1990) Die Ordnung des Lebendigen. Piper-Verlag, München.
Riedl, R.; Krall, P. (1994): Die Evolutionstheorie im wissenschaftstheoretischen Wandel. In: Wieser, W. (Hrsg.): Die Evolution der Evolutionstheorie. Von Darwin zur DNA. Spektrum Akademischer Verlag, Heidelberg, 234-266.
