

Wir haben uns intensiv mit den biochemischen Grundlagen der Photosynthese befasst. Dort lernten wir, dass Pflanzen aus CO2 und Wasser Zucker und Sauerstoff (O2) herstellen können.
Nun wird von einigen „Klima-Querdenkern“ gerne folgende Behauptung aufgestellt:
„CO2 ist kein Schadstoff. Es ist wesentlich für alles Leben auf der Erde. Die Photosynthese ist ein Segen. Mehr CO2 schont die Natur und die Erde: Zusätzliches CO2 in der Luft hat das Wachstum der globalen Pflanzenbiomasse gefördert. Es ist auch gut für die Landwirtschaft und erhöht die Ernteerträge weltweit.“
CO2 ist Pflanzennahrung. Das stimmt. Ist CO2 ein Schadstoff?
Das kommt auf die Konzentration an. Für Pflanzen ist CO2 Nahrung, würde die Atmosphäre aber nur aus CO2 bestehen, würden wir nicht überleben können. Schadstoffe sind immer eine Frage der Konzentration. Daher ist die Frage berechtigt: Ist mehr CO2 automatisch gut (für Pflanzen)?
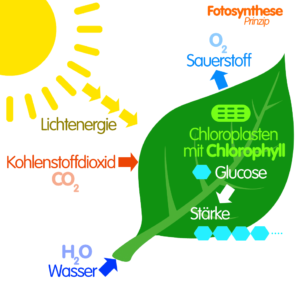
Abb. 1: Photosynthese
Vielleicht hilft ein Vergleich: Wir Menschen müssen Nährstoffe aufnehmen in Form von Kohlenhydraten, Proteinen und Fetten. Ein wichtiger Kohlenhydrat-Lieferant ist Saccharose, unser Haushaltszucker. Er schmeckt nicht nur gut, sondern liefert auch Energie. Aber was passiert, wenn wir zu viel Zucker zunehmen? Wir leiden an Karies, Adipositas, Herz-Kreislauf-Erkrankungen und Diabetes. Ist zu viel Zucker gut für uns? Nein. Warum muss also mehr CO2 automatisch gut für (alle) Pflanzen sein?
Pflanzen leben nicht von CO2 alleine. Damit Pflanzen optimal wachsen können braucht sie auch für sie entsprechende Temperatur- und Feuchtigkeitsbedingungen.
Hier kommt es natürlich auch auf die Pflanzenart an. So wie der Mensch kein Regenwurm ist, ist eine Eiche kein Kaktus. Jede Pflanzenart reagiert unterschiedlich auf das Klima. Manche kommen mit Trockenheit besser klar als andere. Das hängt von vielen Faktoren ab. Die globale Erwärmung sorgt dafür, dass gerade die Temperatur- und Niederschlagsbedingungen sich so ungünstig auf das Wachstum vieler Kulturpflanzen auswirken, dass durch die erhöhte CO2-Konzentration nicht kompensiert werden kann (vgl. Lobell et al. 2011, Zhao & Running 2010).
Die meisten wissenschaftlichen Studien zur CO2-Steigerung wurden bisher nur in solchen geschlossenen Gewächshäusern oder in einzelnen Wachstumskammern durchgeführt. Erst vor kurzem haben Forscher begonnen ihre Aufmerksamkeit auf Experimente im Freien zu lenken. Diese Free-Air CO2 Enrichment- oder “FACE”-Studien untersuchen die Pflanzen im Freiland, wenn sie erhöhten CO2-Konzentrationen ausgesetzt sind. Die Ergebnisse dieser Studien sind bei weitem nicht so vielversprechend wie die von Gewächshausstudien. Die endgültigen Ertragswerte liegen in den Freiluftstudien im Vergleich zu Gewächshausstudien durchschnittlich um 50% niedriger (Leaky et al. 2009, Long et al. 2006, Ainsworth 2005). Gründe für das bessere Wachstum in Gewächshäusern liegen z. B. darin, dass Pflanzen in Gewächshäusern isolierter voneinander sind, das Wurzelwachstum regelmäßiger ist und Schädlinge nicht so leicht eindringen können.
Abgesehen hiervon spielen noch weitere Faktoren eine Rolle: Verschiedene Pflanzenarten reagieren auf erhöhte CO2-Konzentrationen auf unterschiedliche Weise. Sie sind abhängig von der Pflanzenart, aber auch Alter, genetischer Variation, Photosyntheseleistung, Jahreszeit, atmosphärische Zusammensetzung, konkurrierende Pflanzen, Krankheits- und Schädlingsbefall, Feuchtigkeitsgehalt, Nährstoffverfügbarkeit, Temperatur und Verfügbarkeit von Sonnenlicht. Die fortgesetzte Zunahme von CO2 wird ein starkes Treibmittel für eine Vielzahl von Veränderungen darstellen, die für den Erfolg vieler Pflanzen entscheidend sind, die natürlichen Ökosysteme beeinflussen und große Auswirkungen auf die globale Lebensmittelproduktion haben. Der weltweite Anstieg von CO2 ist somit ein großes biologisches Experiment mit unzähligen Komplikationen, die es sehr schwierig machen, den Nettoeffekt dieses Anstiegs mit einem nennenswerten Detailgrad vorherzusagen.
Im Zusammenhang mit der Photosynthese spielt aber ein weiterer Faktor eine Rolle, der auch zeigt, dass Photsynthese nicht gleich Photosynthese ist: Die Photorespiration oder auch Lichtatmung genannt.
Photorespiration
Mit der Lichtreaktion kann die Pflanze das Sonnenlicht sammeln und die Lichtenergie in chemische Energie, gebunden als ATP, speichern. Diese wird bei der Dunkelreaktion, also dem Calvin-Zyklus, genutzt, um Kohlenhydrate herzustellen.
Beim Calvin-Zyklus spielt das Enzym Ribulose-1,5-bisphosphat-Carboxylase-Oxygenase, kurz RuBisCo, eine Schlüsselrolle (Abb. 2). Es fixiert das CO2 aus der Luft auf den C5-Zucker Ribulose-1,5-bisphosphat (RubP). Durch weitere Umwandlungsschritte kann daraus Glucose oder andere C6-Zucker hergestellt werden.
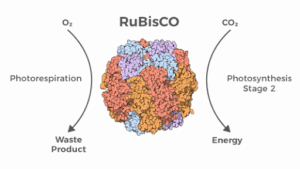
Abb. 2: Ribulose-1,5-bisphosphat-Carboxylase-Oxygenase, kurz RuBisCo
RuBisCo hat allerdings auch noch eine weitere Eigenschaft, die man sich auch durch ihnen Namen herleiten kann. Neben der Kohlenstoffdioxidfixierung (=Carboxylierung) kann es auch Sauerstoff fixieren (=Oxygenierung). Dadurch geht dem Calvin-Zyklus dann quasi ein Kohlenstoffatom „verloren“. Fixiert RuBisCo statt CO2 Sauerstoff, entsteht ein C2-Molekül, das 2-Phosphoglycolat.
Dieses muss in einem energieaufwendigen Weg in verschiedenen Zellorganellen (Mitochondrien und Peroxisomen) zu 3-Phosphoglycerinsäure umgewandelt werden. 3-Phosphoglycerinsäure ist das zwischenprodukt des Calvin-Zyklus, dass nach der Fixierung von CO2 entsteht. Denn nur dieses Molekül kann der Calvin-Zyklus weiter verstoffwechseln. Das C2-Molekül 2-Phophogylcerat ist für den Calvin-Zyklus unbrauchbar, man kann mit ihm also kein Zucker herstellen.
Den Weg zur Wiederverwertung des Kohlenstoffgerüstes wird auch als Photorespiration oder Lichtatmung bezeichnet (Abb. 3). Da er ziemlich viel Energie „verschwendet“, versuchen ihn Pflanzen nach Möglichkeit zu vermeiden.
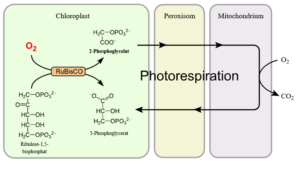
Abb. 3: Photorespiration
Am idealsten ist, wenn RuBisCo möglichst wenig mit Sauerstoff in Berührung kommt und eher das CO2 fixiert. Nun könnte man einwenden: Hey dann ist mehr CO2 in der Atmosphäre doch gut für die Pflanzen. Nicht unbedingt. Mehr CO2 in der Luft sorgt für höhere Temperaturen und weniger Luftfeuchtigkeit.
Besonders an heißen, trockenen Tagen kommt es häufig zur Lichtatmung, da die Pflanzen ihre Spaltöffnungen schließen, damit kein Wasser verdunsten kann. Dadurch kann aber auch wenig Kohlenstoffdioxid aus der Luft in die Pflanzen gelangen und der Sauerstoffgehalt steigt im Vergleich zum Kohlenstoffdioxidgehalt in der Pflanze. Das wiederum führt dazu, dass das Enzym RubisCO auch öfter Sauerstoff an das Akzeptormolekül bindet, die Lichtatmung also stattfindet.
Also bei Trockenheit geschieht es, dass die Spaltöffnungen der Pflanzen sich schließen, sich in der Pflanze der Sauerstoff ansammelt und immer weniger CO2 aufgenommen wird und das Enzym RuBisCO nutzt dann eher den Sauerstoff zur Bildung von 2-Phosphoglycolat, also dem C2-Zucker. Da dieser für den Calvinzyklus „nutzlos“ ist, wird er unter Energieverbrauch in 3-Phosphoglycerinsäure umgewandelt und in den Calvinzyklus zur Produktion von Zucker wieder integriert. Sind die Temperaturverhältnisse ungünstig hoch bleiben die Spaltöffnungen länger geschlossen, was dann dazu führen kann, dass die Pflanzen mehr Energie verbrauchen als sie erzeugen – oder anders ausgedrückt: die Pflanze stirbt ab! Wie lange Pflanzen die Lichtatmung verwenden können ist natürlich von Art zu Art unterschiedlich.
Photosynthese Typen
Es gibt aber im Pflanzenreich verschiedene Strategien die Probleme der Lichtatmung zu umgehen. Es gibt nämlich verschiedene Photosynthese-Typen unterscheiden: die C3– Pflanzen, die C4– Pflanzen und die CAM-Pflanzen (Abb. 4).
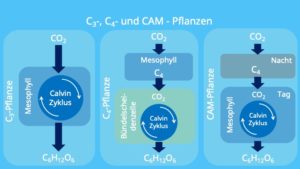
Abb. 4: C3, C4 und CAM Pflanzen
C3 Pflanzen
C3– Pflanzen sind die ganz „normalen Pflanzen“ ohne besondere Anpassungen hinsichtlich der Vermeidung zur Photorespiration verstehen. Das bedeutet, dass der Calvin-Zyklus, so wie wir ihn im Kapitel zur Photosynthese beschrieben haben, abläuft.
Der Name C3 kommt daher, da das erste Produkt nach der Kohlenstofffixierung, also der Anlagerung von Kohlenstoffdioxid an das Akzeptormolekül (1,5-Ribulosebisphosphat) im Calvin Zyklus, ein C3 Körper (3-PGS) ist.
Die meisten Pflanzen auf der Erde betreiben diese Art der Photosynthese, darunter alle Bäume und auch Soja- oder Reispflanzen.
C4 Pflanzen
Die C4– Pflanzen haben jetzt eine Strategie entwickelt, um dem verschwenderischen Weg der Lichtatmung zu entgehen. Ziel dieser Strategie ist es, dass das Enzym RuBisCO immer einer hohen Konzentration an Kohlenstoffdioxid ausgesetzt ist, damit es nicht die Möglichkeit hat, an Sauerstoff zu binden (Abb. 5 und 6).
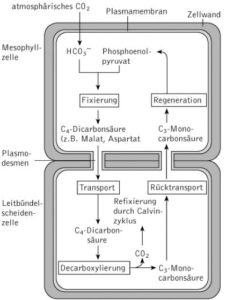
Abb. 5: C4-Photosynthese. 1. CO2 wird vorfixiert. 2. Die daraus gebildete C4-Verbindung wird in das benachbarte Kompartiment transportiert. 3. Dort wird CO2 freigesetzt und im Calvin-Zyklus verbraucht. 4. Der übrige C3-Körper wird wieder zurücktransportiert und für einen weiteren Zyklus regeneriert.

Abb. 6: typische C4-Pflanzen
Die spezielle Anpassung der C4– Pflanzen ist eine räumliche Trennung zwischen der Kohlenstoffdioxid-Fixierung und dem eigentlichen Calvin-Zyklus.
Es findet hier nämlich eine Vorfixierung von Kohlenstoffdioxid statt. In den sogenannten Mesophyllzellen wird das CO2 mit dem Phosphoenolpyruvat (PEP), ein energiereiches Stoffwechselzwischenprodukt der Glykolyse, fixiert, sodass als Zwischenprodukt Oxalacetat entsteht. Oxalacetat ist eine organische Säure mit 4 Kohlenstoffatomen. Daher kommt auch der Name C4– Pflanzen zustande. Oxalacetat wird dann Dicarbonsäure wie Malat oder Asparat umgewandelt.
Die entstehende Säure kann dann weiter in spezielle Zellen (Bündelscheidenzellen) transportiert werden, die sich wie eine Art Kranz anordnen. Dort findet die Kohlenstoffdioxidspaltung der Säure statt und der Calvin-Zyklus kann dann wie gewohnt ablaufen.
Etwa 3% der Arten der Bedecktsamer betreiben eine C4-Fotosynthese und machen etwa 5% der Biomasse aus. Sie fixieren aber 23% des CO2 (Bond et al. 2005, Kellogg 2013). Hierzu gehören vornehmlich Gräser, die aus wärmeren Gebieten mit längeren Trockenperioden stammen, z. B. Mais, Zuckerrohr oder Hirse (Abb. 6). C4-Photosynthese kommt aber auch in anderen Pflanzenfamilien vor und ist wahrscheinlich 65-mal unabhängig voneinander in 19 Pflanzenfamilien entstanden. Bei Bäumen kommt die C4-Photosynthese mit wenigen Ausnahmen praktisch gar nicht vor (Sage 2017, Bresinski et al. 2008, S. 312).
Gegenüber C3-Pflanzen zeichnen sich C4-Pflanzen bei Wasserknappheit, hohen Temperaturen und Sonneneinstrahlung aus. C4-Pflanzen dominieren also eher trockene Gebiete, wie wir sie z. B. aus den Savannen Afrika oder Australien kennen (Abb. 7). So betreiben etwa 70 % aller im Death-Valley-Nationalpark lebenden Arten eine C4-Photosynthese (Bresinski et al. 2008, S. 311).
In tropischen Regenwäldern dominieren die C3-Pflanzen (Abb. 7), was nicht verwundern sollte, da die C4-Photosynthese in Bäumen praktisch gar nicht vorkommt. Bei Nadelbäumen, Farnen und Moosen kommt eine C4-Photosynthese gar nicht vor (Sage 2017).

CAM Pflanzen
Auch die CAM-Pflanzen (=Crassulaceen-Säurestoffwechsel) besitzen eine spezielle Strategie, um die Lichtatmung zu minimieren. Hier erfolgt eine zeitliche Trennung zwischen der Kohlenstoffdioxidfixierung und dem Ablauf des Calvin Zyklus (Abb. 8).
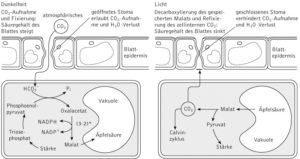
Abb. 8: CO2-Fixierung bei CAM-Pflanzen
Das läuft so ab: In der Nacht öffnen die Pflanzen ihre Spaltöffnungen, damit CO2 in die Blätter diffundieren kann. Dort kann Kohlenstoffdioxid in eine organische Säure mit eingebaut werden (ähnlich zu den C4 Pflanzen). Die dadurch entstehende Säure aus 4 Kohlenstoffatomen (meistens Äpfelsäure) kann dann in die Vakuolen der Zellen transportiert und dort gespeichert werden.
Tagsüber schließen die Pflanzen wieder ihre Spaltöffnungen, um den Wasserverlust zu verringern. Die Säure kann nun aus den Vakuolen transportiert und Kohlenstoffdioxid abgespalten werden. Dadurch kann der Calvin-Zyklus wieder wie gewohnt ablaufen. Auch hier ist Ziel dieser Strategie, dass das Enzym RuBisCO immer einer hohen Konzentration an Kohlenstoffdioxid ausgesetzt ist, damit es nicht die Möglichkeit hat, an Sauerstoff zu binden.
Zu den CAM-Pflanzen gehören nicht nur die sukkulenten Dickblattgewächse (Crassulaceae), nach denen dieser Typ der CO2-Fixierung benannt ist, sondern auch viele Arten aus den Familien Cactaceae, Agavaceae und Euphorbiaceae. Die Ananas ist ebenfalls eine CAM-Pflanze. Eine Reihe von Pflanzen sind zudem in der Lage, den CAM-Stoffwechsel fakultativ zu betreiben. So wechselt die Eispflanze, Mesembryanthemum crystallinum (Aizoaceae), bei Wassermangel vom C3-Weg (C3-Pflanzen) zum CAM-Weg. CAM-Pflanzen kommen vorwiegend in Trockengebieten vor, sind aber auch in tropischen Regenwäldern zu finden. Denn viele Aufsitzerpflanzen, sog. Epiphyten, also Pflanzen, die auf anderen Pflanzen wachsen, leiden auch unter Wassermangel und hohen Temperaturen. Hierzu zählen z. B. viele Orchideen und Bromelien. Entsprechend kommt auch bei ihnen die CAM-Photosynthese relativ häufig vor (Abb. 9).

Abb. 9: typische CAM-Pflanzen
Abhängigkeit von der CO2-Konzentration
Der Kurvenverlauf für die Abhängigkeit der Fotosyntheseleistung von der CO2-Konzentration entspricht einer Sättigungskurve (Abb. 10). Das heißt zu einem bestimmten Zeitpunkt wird ein Maximum der Photosynthese-Leistung erreicht und die weitere Zugabe von CO2 verstärkt die Photosynthese-Leistung nicht.
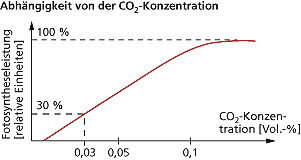
Abb. 10: Abhängigkeit der Photosyntheseleistung von der CO2-Konzentration.
Die Photosynthese-Leistung steigt bei den meisten Pflanzen proportional mit der Erhöhung des CO2-anteils bis auf ca. 0,1 Vol.-%. Danach folgt bei vielen Pflanzen trotz steigender CO2-konzentration keine Zunahme der Photosynthese-Leistung (CO2-sättigung) mehr, wobei ein zu hoher Kohlenstoffdioxidanteil auch zu Schädigungen führen kann (hieraus erkennt man schon, dass eine ewige Steigerung des CO2 in der Atmosphäre nicht unbedingt zum Pflanzenwachstum führen muss, denn irgendwann ist auch eine Pflanze satt).
Der CO2-Kompensationspunkt gibt an, ab welchem Punkt die CO2-Aufnahme und CO2-Abgabe, also die Leistung von Photosynthese und Atmung in einer Pflanze, im Gleichgewicht sind. Bei C3-Pflanzen liegt dieser – je nach Art – zwischen 0,005 und 0,01 Vol.-% CO2 in der Atmosphäre.
Unter natürlichen Bedingungen ist bei C3-Pflanzen der CO2-Anteil in der Atmosphäre (0,03 Vol.-%) der begrenzende Faktor für die Photosynthese-Leistung. Die Photosynthese-Intensität kann bei diesen Pflanzen durch CO2-Anreicherung in Gewächshäusern bis zum Dreifachen gesteigert werden. Wir haben aber schon vorhin kennengelernt, dass die Situation in der freien Natur eine andere ist, als in Gewächshäusern.
Photosynthese-Spezialisten wie C4- und CAM-Pflanzen erhöhen in Anpassung an sonnige trockene Standorte unter Energieverbrauch die Konzentration an CO2, indem sie zunächst einen anderen Akzeptor (nämlich das Phosphoenolpyruvat PEP) zur CO2-Fixierung verwenden. So können sie auch bei sehr niedrigen CO2-Konzentrationen noch eine hohe Photosynthese-Leistung erzielen.
Weitere Photosynthese-Faktoren
Chlorophyllgehalt
Der Chlorophyllgehalt einer Pflanze ist unter natürlichen Bedingungen kein begrenzender Faktor. So können selbst Pflanzen mit geringem Chlorophyllgehalt ausreichend Photosynthese betreiben. Der Chlorophyllgehalt kann aber bei Pflanzen in Anpassung an unterschiedliche Lichtintensitäten variieren. So weisen Schattenblätter im Vergleich zu Sonnenblättern einen höheren Chlorophyllgehalt pro Einheit Blattfläche auf (Abb. 11).
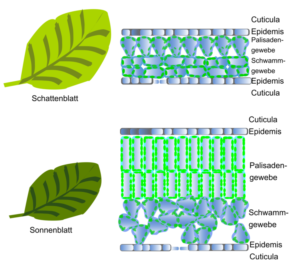
Abb. 11: Unterschied Sonnenblatt, Schattenblatt.
Licht
Die Photosyntheseleistung hängt natürlich von der Lichtmenge und -intensität ab, weil die für den Calvinzyklus notwendigen ATP und NADPH-Moleküle in der Lichtreaktion erzeugt werden.
Abb. 12 zeigt die Abhängigkeit der Photosyntheseleistung von der Lichtintensität:
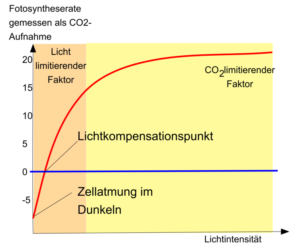
Abb. 12: Abhängigkeit der Photosyntheseleistung von der Lichtintensität
Der Kurvenverlauf für die Abhängigkeit der Fotosyntheseleistung von der Lichtenergie beginnt im negativen Bereich, da hier vorerst der Sauerstoffverbrauch durch die Zellatmung größer ist als die Sauerstoffproduktion durch die Fotosynthese. Solange keine anderen Umweltfaktoren (z. B. Kohlenstoffdioxid, Temperatur) begrenzend wirken, wächst zunächst die Fotosyntheseintensität mit Erhöhung der Lichtintensität proportional. Am Lichtkompensationspunkt, wo die Sauerstoffabgabe (bzw. Kohlenstoffdioxidaufnahme) durch die Fotosynthese genauso so groß ist wie der Sauerstoffverbrauch (bzw. Kohlenstoffdioxidproduktion), schneidet die Kurve die x-Achse und die Nettofotosynthese hat den Wert null.
Nur Pflanzen, deren Produktion von Sauerstoff bzw. Verbrauch an Kohlenstoffdioxid über dem Lichtkompensationspunkt liegt (höhere Fotosyntheseleistung als Atmungsintensität), können organische Stoffe speichern. Mit zunehmender Strahlungsintensität wirken die anderen Faktoren nach dem Gesetz des Minimums immer stärker limitierend auf die Fotosyntheseleistung. Dadurch steigt die Kurve der Fotosyntheseleistung immer weniger und zwar bis zu dem Punkt, an dem sich trotz steigender Lichtintensität die Fotosyntheseleistung nicht mehr erhöht (Lichtsättigung).
Der Kurvenverlauf der Fotosyntheseintensität in Abhängigkeit von der Lichtintensität wird dementsprechend als Sättigungskurve bezeichnet. Falls die Lichteinwirkung trotzdem weiter ansteigt, kann dies zu einer Schädigung der Fotosysteme führen. Es werden nämlich sog. Sauerstoffradikale (O2–) gebildet. Als Radikale bezeichnet man Atome oder Moleküle mit mindestens einem ungepaarten Elektron, die meist besonders reaktionsfreudig sind. Sauerstoffradikale sind schädliche Formen des Sauerstoffs, die auch ein Nebenprodukt der Atmungskette der Mitochondrien sind. Werden zu viele dieser radikale gebildet, spricht man vom oxidativen Stress und sie spielen bei verschiedensten Erkrankungen sowie beim Alterungsprozess eine wesentliche pathophysiologische Rolle. Z. B. greifen sie Proteine oder die DNS an und zerstören sie.
Unter natürlichen Bedingungen treten bei Pflanzen Sauerstoffradikale auf, z. B. bei sehr geringen Temperaturen oder bei Schattenpflanzen, wenn sie plötzlich sehr starkem Lichteinfluss ausgesetzt werden, auf. In begrenztem Umfang können bestimmte Fotosynthesepigmente (Xanthophylle) überschüssige, nicht verwertbare Lichtenergie zum Schutz vor entstehenden Sauerstoffradikalen auffangen.
Pflanzenarten zeigen unterschiedliche Anpassungen an die Lichtintensität. So unterscheiden sich Fotosynthesespezialisten, wie der Mais als C4-Pflanze, von den C3-Pflanzen. C4-Pflanzen erreichen selbst bei hoher Lichtintensität keine absolute Lichtsättigung. Deshalb sind C4-Pflanzen den C3-Pflanzen bei voller Sonneneinstrahlung in Bezug auf die Strahlungsverwertung überlegen. Bei C3-Pflanzen unterscheidet man an Starklicht (Sonne) und Schwachlicht (Schatten) angepasste Pflanzen oder Organe.
Temperatur
Wie bei allen enzymatisch gesteuerten Reaktionen ist auch die Fotosyntheseleistung von der Temperatur abhängig und wird in Form einer Optimumskurve beschrieben (Abb. 13).
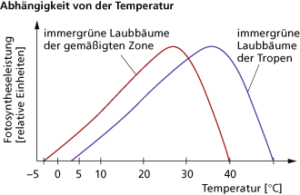
Abb. 13: Abhängigkeit der Photosynthese von der Temperatur
So steigt die Fotosyntheseleistung bei ausreichender Versorgung mit den anderen Bedingungen entsprechend der RGT-Regel bis zum Optimum (RGT-Regel: Erhöhung der Temperatur um 10 K bewirkt eine Leistungssteigerung auf ca. das Doppelte). Nach Erreichen des Optimums wirken andere Faktoren immer stärker limitierend auf die Fotosyntheseleistung, so u. a. Licht- und Kohlenstoffdioxidversorgung, Erhöhung der Atmungsintensität oder Denaturierung der beteiligten Enzyme. Gleichzeitig haben höhere Temperaturen Einfluss auf den Wassergehalt innerhalb der Pflanze. Durch die erhöhte Transpiration sinkt der Wassergehalt in der Pflanze, sodass sich bei höheren Temperaturen die Spaltöffnungen schließen und so die Kohlenstoffdioxidzufuhr stark behindern. Die Werte der Optimumkurve können innerhalb der Pflanzenart und zwischen den Arten sehr unterschiedlich sein und können durch Temperaturanpassungen verschoben werden.
Wasser und Mineralstoffe
Wasser ist neben Kohlenstoffdioxid ein direkter Ausgangsstoff für die Fotosynthese. Die benötigte Menge an Wasser ist jedoch so gering, dass sie sich nicht direkt auf den Stoff- und Energiewechselprozess auswirkt. Vielmehr werden indirekt durch den Wassermangel die enzymatischen Prozesse gehemmt und durch den Verschluss der Spaltöffnungen wird die Kohlenstoffdioxidaufnahme entscheidend beeinträchtigt. Die Pflanzen greifen zunächst auf intern entstandenes Kohlenstoffdioxid der Zellatmung zurück, bevor in weiteren Stufen die Kohlenstoffdioxidzufuhr ganz gedrosselt wird und dann Schäden an Fotosyntheseapparat und Membranen auftreten.
Die Mineralstoffe, die über die Wurzel in die Pflanze aufgenommen werden, sind u. a. für den Ablauf aller biochemischen Prozesse und auch zum Aufbau von Zellstrukturen notwendig. Chloroplasten benötigen Mineralstoffe (Mg, Fe, Cu, Mn) für den Bau des Fotosyntheseapparats. Diese sind außerdem Bestandteil der Pigmente und Redoxsysteme (Elektronentransportkette).
Zusammenfassend können wir sagen: Pflanzenwachstum hängt nicht alleine von CO2 ab, sondern ist etwas komplexer. Das sollte man immer bedenken, wenn man Aussagen hört wie CO2 sei gut für das Pflanzenwachstum.
Literatur
Ainsworth, E. A., Long, S. P. (2005): What have we learned from 15 years of free-air CO2 enrichment (FACE)? A meta-analytic review of the responses of photosynthesis, canopy properties and plant production to rising CO2. New Phytol.;165(2):351-71. doi: 10.1111/j.1469-8137.2004.01224.x. PMID: 15720649.
Bond, J., Woodward,, F. I., Midgley, G. F. (2005): The global distribution of ecosystems in a world without fire. New Phytologist. 165, Nr. 2, S. 525–538. doi:10.1111/j.1469-8137.2004.01252.x. PMID 15720663.
Bresinsky, A. et al. (2008): Strasburger – Lehrbuch der Botanik. 36. Auflage. Spektrum Akademischer Verlag; Heidelberg; ISBN 978-3-8274-1455-7, S. 312.
Kellogg, E. A. (2013): C4 photosynthesis. In: Current Biology. 23, Nr. 14, S. R594–R599. doi:10.1016/j.cub.2013.04.066.
Leakey, A. D. B., Ainsworth, E. A., Bernacchi, C. J., et al. (2009): Elevated CO2 effects on plant carbon, nitrogen, and water relations: six important lessons from FACE, Journal of Experimental Botany, Volume 60, Issue 10, 2859–2876, https://doi.org/10.1093/jxb/erp096
Lobell, D., Bänziger, M., Magorokosho, C., et al. (2011): Nonlinear heat effects on African maize as evidenced by historical yield trials. Nature Clim Change 1, 42–45. https://doi.org/10.1038/nclimate1043
Sage, R. F. (2017): A portrait of the C4 photosynthetic family on the 50th anniversary of its discovery: species number, evolutionary lineages, and Hall of Fame. Journal of Experimental Botany. Band 68, Nr. 2, S. e11–e28, doi:10.1093/jxb/erx005, PMID 28110278.
Zhao, M., Running, S. W. (2010): Drought-Induced Reduction in Global Terrestrial Net Primary Production from 2000 Through 2009. Science Vol. 329, Issue 5994, pp. 940-943 DOI: 10.1126/science.1192666
