
Eine Zusammenstellung von Michael Kubi (2018)
als pdf: Organismen als hydraulische Konstruktionen
Morphologie – was ist das?
Die Morphologie (Gestaltlehre) ist eine der grundlegenden Wissenschaftszweige in der Biologie. Denn sie erforscht den Bau und die Funktionen eines Organismus. Durch sie werden z. B. die Baupläne im Tierreich erkannt. In einem Bauplan werden die gemeinsamen neuen und ursprünglichen Merkmale einer systematischen Gruppe (Taxon) dargestellt. Im Tierreich werden gegenwärtig 36 Grundbaupläne (BURDA et al. 2008, siehe Abb.1) unterschieden, aus denen sich alle Lebewesen entwickelt haben und im Laufe der Evolution differenzierten. Diese Grundbaupläne werden als Stämme oder Phylum bezeichnet. Zu den bekanntesten Stämmen im Tierreich gehören z. B. die Chordaten (wozu auch die Wirbeltiere gehören), Arthropoden (Insekten, Spinnen, Krebse, Tausendfüßer), Anneliden (Regenwürmer) und Mollusken (Schnecken, Muscheln, Tintenfische).
Ziel der Morphologie ist es dabei, nicht nur den Bauplan der Organismen zu beschreiben, sondern auch die Verwandtschaftsbeziehungen zu ermitteln, um so die Evolutionsgeschichte zu rekonstruieren. dabei arbeitet die Morphologie vergleichend:
„Erst durch den Vergleich wird Morphologie zur Wissenschaft (…). Erst das ‚In-Beziehung-setzen‘ neugewonnener mit alten Daten macht Morphologie zur Wissenschaft von den gestalten, zur ‚Gestaltenlehre‘. Grundsätzlich gilt, dass überhaupt nur Ähnlichkeiten erkannt werden können; alle anderen Schlussfolgerungen auf der Basis solcher erkannten Ähnlichkeiten sind als Interpretationen mit Wahrscheinlichkeitscharakter anzusehen. Denn Evolution ist ein einzigartiger, prinzipiell nicht wiederholbarer Prozess, der sich der direkten Analyse entzieht und nur Rekonstruktionen erlaubt. Damit steht die Biologie übrigens nicht alleine da: Alle historischen Disziplinen, wie Kosmologie, Archäologie oder Mittelalterliche Heraldik, haben dasselbe Problem.“ (Haszprunar 1994)
Die Morphologie arbeitet entweder funktionell (Funktionsmorphologie), die sich mit der Funktionsweise der Lebenserscheinungen der Organismus befasst oder historisch, die versucht einen allgemeinen Bauplan sämtlicher Organismengruppen zu ermitteln. Letztere wurzelt auf der sogenannten idealistischen Morphologie (Haszprunar 1994).
Unter idealistischer Morphologie versteht man eine von J. W. Goethe entwickelte Methode eine Ordnung in der Vielfalt der Organismen zu schaffen. Dabei werden zwischen den Organismen und ihren Strukturen bestehende Ähnlichkeiten auf einen gemeinsamen Urtyp (Archetypus) zurückgeführt. Die idealistische Morphologie wurzelt in der Ideenlehre Platons, für den die realen Phänomene unvollkommene Widerspiegelungen einiger Ideen, die unabhängig von realen Objekten bestehen. Abwandlungen vom Archetypus wurden von Goethe als Metamorphosen bezeichnet. In der Evolutionsforschung werden diese „Archetypen“ jedoch anders als bei Goethe nicht als ideelle Gebilde, sondern als real existierende Lebewesen aufgefasst, die sich wie die heutigen Formen an die Umwelt anpassen.
Folgende Definition findet sich für den Begriff der idealistischen Morphologie:
„[E]ine vor dem Aufkommen der Evolutionstheorie vor allem von J.W. von Goethe entwickelte Methode, die in der Mannigfaltigkeit der Organismen herrschende ‚Ordnung‘ zu erfassen und darzustellen. In der idealistischen Morphologie werden die zwischen verschiedenen Organismen und ihren Strukturen bestehenden ‚typischen Ähnlichkeiten‘ auf einen gemeinsamen Typus (Archetypus) zurückgeführt, eine ‚Urform‘ oder ein ‚Urbild‘. (…) Die idealistische Morphologie wurzelt daher in der Ideenlehre Platons, für den die veränderlichen realen Phänomene nichts anderes als (unvollkommene) Widerspiegelungen einer begrenzten Anzahl von ‚Ideen‘ (eideai) oder Wesenheiten (Essenzen) waren, die unabhängig von den realen Objekten bestehen. Die stufenweisen (durch ‚Übergänge‘ verbundenen) ‚Abweichungen‘ der realen Formen vom Typus werden von Goethe als ‚Metamorphosen‘ aufgefaßt, vergleichbar den Variationen eines musikalischen Themas. Gemäß dieser Vorstellung entwarf Goethe eine ‚Urpflanze‘ (Goethe, Abb.), als ein ‚Modell‘, von dem sich alle Blütenpflanzen ableiten lassen (‚Metamorphose der Pflanze‘, 1790). In gleicher Weise bemühte er sich um die Aufstellung eines ‚osteologischen Typus‘ der Wirbeltiere (genauer der Vierfüßer) (‚Versuch über die Gestalt der Tiere‘, 1790). (…) Die idealistische Morphologie hat wesentliche Grundlagen für die vergleichende Morphologie (der Begriff wurde von Goethe 1795 in die Wissenschaft eingeführt) und für ein ‚typologisches System‘ der Organismen geschaffen und damit gleichzeitig wichtige Voraussetzungen für die Entwicklung des Evolutionsgedankens durch J. Lamarck und C.R. Darwin. Die Evolutionstheorie erklärt heute die Übereinstimmungen (Homologien) im ‚Bauplan‘ verschiedener Organismen als gemeinsames Erbe von einem gemeinsamen Ahnen. (…) Im Gegensatz zum ‚Urbild‘ der idealistischen Morphologie war der gemeinsame Ahne einer aus ihm entwickelten Gruppe ein real existierendes Lebewesen, das keine ‚reine‘ Form repräsentieren konnte, sondern dessen Organe selbstverständlich Anpassungen an ihre spezielle Funktion und an die Umweltbedingungen aufweisen mußten. Die Abwandlungen der Organe in der Phylogenese (Phylogenie) sind auch nicht ‚unvollkommene‘ Manifestationen der ihnen zugrundeliegenden Wesenheiten (wie im Essentialismus platonscher Prägung), sondern Ausdruck unterschiedlicher Anpassungen.“ (https://www.spektrum.de/lexikon/biologie/idealistische-morphologie/33644)
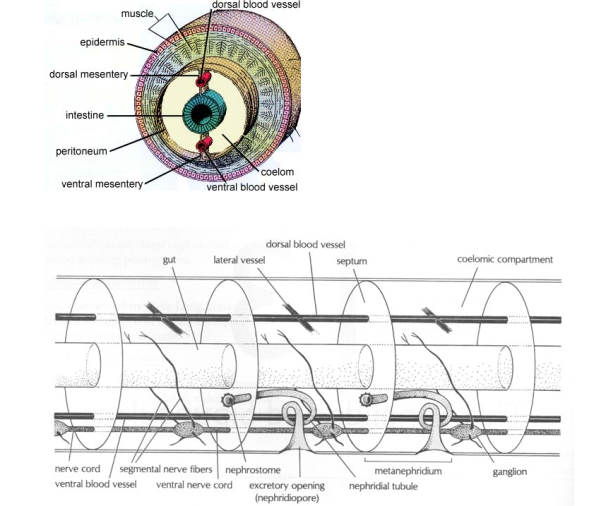
Abb. 1: Bauplan der Anneliden, einer der 36 Baupläne im Tierreich. Quelle: https://projects.ncsu.edu/project/bio402_315/Annelids/annelid.html
Nach Haszprunar (1994) war die historische Betrachtungsweise zu Beginn des 20. Jahrhunderts dominierend, da mit den Erkenntnissen Darwins die Ursache der Ähnlichkeiten gemeinsame Abstammung waren. Die ausschließlich historische Betrachtungsweise erlebte jedoch zunehmend einen kontinuierlichen Niedergang, da die funktionelle Morphologie zeigen konnte, dass viele Strukturen zu einem hohen Anteil primär funktionell und nicht historisch zu erklären seien.
In den 1960er Jahren konnte sich zunehmend die vom deutschen Morphologen Willi Henning begründete Phylogenetische Systematik oder auch „Kladistik“ genannt (Henning 1950, 1966) durchsetzen, welches die historische Sichtweise wiederbelebte.
Ein jeder Organismus, will man ihn beschreiben, hat für ihn typische charakteristische Eigenschaften, die man als Merkmale definiert. Diese können Strukturmerkmale sein (also die Anatomie), aber auch physiologische (z. B. Warmblütigkeit), ökologische (wasser- oder landlebend) oder ethologische (Verhaltensweisen). Durch einen Vergleich diverser Merkmale lassen sich die Verwandtschaftsgrade der Organismen ableiten. Gehen die Merkmale auf einen gemeinsamen evolutionären Ursprung zurück, spricht man von Homologien. So sind beispielsweise die Gliedmaßen der Wirbeltiere zueinander homolog, also vom prinzipiell gleichen Grundaufbau, da sie alle auf einen Vorfahren zurückgehen.

Abb. 2: Typischer Stammbaum der Säugetiere, basierend auf Merkmalsanalysen. Quelle: wikipedia
Der Biologe Adolf Remane hatte in den 1950er Jahren die Homologie-Kriterien festgelegt, die sich auch heute noch in den Biologie-Lehrbüchern wiederfinden und von weiteren Autoren verfeinert und ergänzt wurden (Remane 1952, Riedl 1975, Tyler 1988).
Dabei sind nicht alle Merkmale gleichwertig: Man unterscheidet zwischen Plesiomorphien und Apomorphien. Apomorphien sind abgeleitete Merkmale, also welche die im Vergleich zum Vorfahren der jeweils betrachteten Stammlinie neu erworben wurden. Plesiomorphien hingegen sind ursprüngliche Merkmale, die schon die Vorfahren hatten. Wenn wir uns als Beispiel nehmen, so stellen wir fest, dass unser Merkmal „Wirbelsäule“ plesiomorph ist, da eine Wirbelsäule evolutionär älter ist als der Mensch und schon bei den Fischen auftaucht. Die Entwicklung einer „artikulierten Lautsprache“ hingegen ist ein Merkmal, dass uns Menschen eigen ist und uns z. B. von unseren nächsten Verwandten – Schimpanse und Bonobo – abtrennt; es ist also eine Apomorphie.
In einem späteren Artikel wird sich intensiver mit der phylogenetischen Systematik – so heißt das oben beschriebene Wissenschaftsfeld, das aufgrund von Merkmalen evolutionäre Verwandtschaftsbeziehungen aufstellt – mit ihren Vor- wie Nachteilen intensiver befassen.
Aber schon hier wird deutlich, dass anhand solcher Beschreibungen sich gut evolutionäre Verwandtschaftsbeziehungen zwischen den Organismen aufstellen lassen.
Doch auch Haszprunar (1994: 133) gibt zu verstehen, dass durch die Wiederbelebung der historisch arbeitenden Morphologie, die funktionellen Aspekte in Frage gestellt werden, da nur die Merkmalsverteilung eine Rolle spiele.
Es besteht also ein immer wieder kehrender Widerspruch zwischen historischen und funktionellen Analysen der Morphologie. Es gibt jedoch Lösungsansätze, beide Aspekte miteinander zu vereinen. Eine ist die sogenannte Konstruktionsmorphologie oder „Frankfurter Evolutionstheorie“, die ab den 1970er Jahren am Forschungsinstitut Senckenberg in Frankfurt am Main Arbeiten publizierte, die eine neue Sicht des Evolutionsgeschehen präsentieren (Gutmann 1972, 1989, 1995, Gutmann & Bonik 1981). Im Zentrum der Frankfurter Evolutionstheorie stehen konstruktionsmorphologische Untersuchungen der Bau- und Funktionsweise der Lebewesen. Entsprechend wird die „Frankfurter Evolutionstheorie“ auch als Konstruktionsmorphologie bezeichnet. Um Verwechslungen oder gar Vergleiche mit der „Frankfurter Schule“ Theodor Adornos zu vermeiden, wird in Folge der Begriff der Konstruktionsmorphologie verwendet. Das hat u. a. auch damit zu, dass im Gegensatz zum pseudomarxistischen Ansatz der Theorien Adornos, die Konstruktionsmorphologie wertvolle Überlegungen liefert. Zum anderen soll die Bedeutung dieses Zweiges der Morphologie nicht auf ihren geographischen Ursprung reduziert werden, sondern vor allem die inhaltliche Ausarbeitung begründet werden.
Einer der Hauptbegründer der Konstruktionsmorphologie ist Wolfgang Gutmann. Die Konstruktionsmorphologie (von griechisch μορφή, morphé = Gestalt, Form und λόγος, lógos = Wort, Lehre, Vernunft) ist die Lehre von der Struktur, der Form und der Formerzeugung sowie der Funktionsweise der Organismen (GUDO 2002 gibt eine vollständige Liste der Publikationen von GUTMANN). Die Konstruktionsmorphologie begreift den Organismus als durchgehend hydraulisch konstruierte Energiewandler. Morphologische Veränderungen sind nicht allein auf Mutationen der DNA-Sequenz und Epigenese zurückzuführen. Die Körperkonstruktion als Ganzes entscheidet in letzter Instanz darüber, welche Veränderungen von Seiten der Gene und Proteine weiterwirken können. Das Funktionsgefüge der Konstruktion bestimmt, welche molekularen Veränderungen bestehen bleiben.
Dieser Theorie, die Organismen als funktionelle Einheiten und nicht als Merkmalsträger untersucht, gingen Vorarbeiten meist angelsächsischer Autoren wie ALEXANDER (1968, 1985), CHAPMANN (1958, 1962), CLARK (1964), MANTON (1950, 1952a, 1952b, 1961, 1965, 1967, 1972, 1973) und WAINWRIGHT (1970, 1975, 1980) voraus und wurden weiterentwickelt. Der Begriff Konstruktionsmorphologie wurde dabei vom deutschen Entomologen Hermann WEBER (1954, 1958) entwickelt, bei der funktionsmorphologische Untersuchungen in eine den gesamten Organismus betreffende Konstruktion eingeführt werden. Dabei soll die Morphologie nicht nur vergleichend arbeiten, sondern den Organismus in seiner Dynamik (also seiner Ontogenie und Phylogenie) berücksichtigen. Seine Ideen sind von SEILACHER (1970) in die Paläontologie eingeführt worden. Danach wird die Konstruktion eines Organismus von historisch-phylogenetischen, ökologisch-adaptiven und bautechnischen Faktoren beeinflusst. GOULD (2002) und CUBO (2004) greifen dieses Konzept in ihren Publikationen wieder auf.
Die Konstruktionsmorphologie GUTMANNS versteht Lebewesen als funktionelle, operational geschlossene, energiewandelne hydraulische Einheiten.
GUDO (2007: 35) fasst die Grundlagen der Konstruktionsmorphologie wie folgt zusammen:
„Lebewesen werden hinsichtlich ihrer Körperkonstruktionen und ihrer spezifischen Eigenschaften als hydraulische, energiewandelnde Konstruktionen verstanden (Organismusbegriff). Da die Evolution durch die innere Organisation gerichtet wird, ist der Begriff der Anpassung lediglich eine beliebige ex-post-Feststellung. Organismen dringen vielmehr in Lebensräume ein und gestalten sie in entscheidender Weise selbst mit. Evolution ist ein weitgehend interdeterministischer Vorgang. Nur die Bedingungen des evolutionären Wandels lassen sich bestimmen. Der Evolutionsmechanismus ist die kontinuierliche Energiewandlung und Reproduktion, durch die eine ununterbrochene Linie besteht und weiter gegeben wird. Die Körperkonstruktion der Organismen verursacht, bestimmt und richtet die evolutionären Veränderungen, und erlaubt es, bestimmte Lebensräume zu erschließen. Organismen sind keine Objekte, sondern Subjekte der Evolution. Morphologische Veränderungen sind nicht allein auf Mutationen der DNA-Sequenz und Epigenese zurückzuführen. Die Körperkonstruktion als Ganzes entscheidet in letzter Instanz darüber, welche Veränderungen von Seiten der Gene und Proteine weiterwirken können. Das Funktionsgefüge der Konstruktion bestimmt, welche molekularen Veränderungen bestehen bleiben. In der Evolution ist nur möglich, was die innere Organisation der Organismen auf weiteren geordneten Bahnen zulässt. Evolution findet – metaphorisch gesagt – auch im Schlaraffenland statt, weil der Antrieb des Wandels thermodynamisch begründet ist und nicht durch Ressourcenbeschränkung und Umweltwandlung.“
VOGEL (1991: 12) kommentiert:
„Konstruktionsmorphologie ist die Erforschung der Organismen und ihrer Teile als kohärente Systeme und Subsysteme, die statisch und dynamisch (d. h. im Ruhezustand und in Bewegung) physikalischer Prinzipien und Prozessen gehorchen. Mit dieser Forschungsrichtung sollen auch Lizenzen und Limitierungen erkundet werden, die sich aus diesem Systemcharakter für Ontogenie und Evolution ergeben. Gefragt ist dabei auch, wie mechanische und chemische Kräfte wechselseitig aufeinander wirken und in welcher Beziehung Konstruktion, Verhalten und Umwelt zueinander stehen.“
Ziel des vorliegenden Artikels soll es sein, die Grundlagen der Konstruktionsmorphologie sensu GUTMANN zusammenfassend wiederzugeben und auf ihre Aktualität zu überprüfen
Parallelen zur Technik
Lebewesen lassen sich am besten verstehen, wenn Parallelen zur Technik gezogen werden können. Dabei werden Organismen wie technische Modelle oder wie Maschinen betrachtet (vgl. etwa Gutmann 1997), aber nicht mit diesen gleichgesetzt:
„Organismen werden so wie Maschinen als material- und energiezehrende Systeme verstanden, in denen die bekannten Naturgesetze gelten. Sie sind wie Maschinen Energiewandler, die aus verschiedenen Teilsystemen zusammengesetzt sind. Sie erbringen Leistungen, indem sie sich durch ständige Aquisition von Material und Energie am Leben erhalten und Nachkommen produzieren. Organismen unterscheiden sich von den technischen Maschinen in der Eigenschaft, sich selbst mit der notwendigen Energie zu versorgen (…) Es wird keine Identität von Organismen und Maschinen behauptet, sondern lediglich in Übereinstimmung in bestimmten Aspekten genutzt, um Lebewesen in einer technischen Weise zu verstehen und zu beschreiben“ (Gudo 1999: 49).
Desweiteren wachsen Organismen im Gegensatz zu Maschinen und betreiben Evolution, was jedoch ebenfalls mechanische Aktion impliziert (VOGEL 1991).
Außerdem können Organismen nicht wie technische Maschinen „wegen Umbau geschlossen“ werden. Evolutionäre Veränderungen müssen stattfinden, während sich der Organismus „in Betrieb“ befindet, woraus sich Limitationen für den „Umbau“ ergeben. Ein Organismus kann somit nicht auf dem Papier neu entworfen werden. Nicht alle Bestandteile des Organismus können umgebaut werden, soll dieser lebensfähig bleiben. Veränderliche Bestandteile werden dabei als Varianzen, Unveränderliche als Invarianzen bezeichnet. Bei der Rekonstruktion der Evolution muss man sich also fragen, wie ein ständig im Lebensbetrieb befindlicher Organismus verändert werden kann, ohne dass es zu katastrophalen Entgleisungen kommt. Ein Phylogenetiker muss folglich ermitteln, welche Konstruktionsteile sich ohne Beeinträchtigung von Funktionsfähigkeit und Fortpflanzung abwandeln lassen. Dabei muss jedes Zwischenstadium voll funktionsfähig sein (GUTMANN 1990, 1997, GUTMANN & BONIK 1981, GUDO et al. 2009, VOGEL 1991).
Organismus und Hydraulik
Eines der wesentlichen Konstruktionsprinzipien von Organismen ist die Hydraulik (Gutmann 1988). Da Stoffwechselvorgänge des Lebens nur im wässrigen Medium stattfinden können, daher aber nicht ins Endlose zerfließen dürfen, besteht der Körper der Organismen aus flexiblen Hüllen und darin eingeschlossener Füllung. Bei dem physikalischen Prinzip der Hydraulik handelt es sich um die Eigenschaft, dass Flüssigkeiten inkompressibel sind. Auch Zellen, die ebenfalls hydraulische Gebilde sind, besitzen eine automatische Tendenz eine Kugelform anzunehmen (GUTMANN 1972a, 1972b, 1983 GRASSHOFF 1993, GUDO 1999). Bei Organismen spricht man von Weichkörper-Hydraulik (GRASSHOFF 1993, GRASSHOFF, GUTMANN & SCHÄFER 1994) und beschreibt dabei den Unterschied zur menschlichen Technik:
„Hydraulik oder Hydroskelett gilt im Bereich des Lebens allerdings in einem Sinne, der von der menschlichen Technik abweicht. Technische Hydraulik ist an feste Röhren und Zylinder gebunden. Die Hydraulik der lebenden Formen liegt in einer völlig flexiblen Membran als total verformbares Gebilde vor.“ (Gutmann 1983)
Die entsprechende Kombination von flexibler Hülle und flüssiger Füllung hat aber auch in technischen Gegenständen, wie Ballons, Kissen und Sandsäcken, ihr Gegenstück. In den Arbeiten von OTTO (1977, 1978, 1979) wird dieses Phänomen als Pneu bezeichnet. Die Weichkörper-Hydraulik der Organismen stellt damit ein Sonderfall des Pneus dar (GRASSHOFF, GUTMANN & SCHÄFER 1994). Hydraulik ist somit die Grundlage der Morphologie (BONIK et al. 1978).
Die generelle Tendenz zu Kugelform wird im Organismus auf verschiedene Weise vermieden. Erst durch Versteifungen, Bandagierungen, interne Verspannungen oder Hüllenvorformierungen lassen sich andere Gestalten erzeugen (GUDO 1999, Abb.3 Kugelform). Unverspannte Bereiche nehmen immer eine runde Form an, da dies energetisch am günstigsten ist (GUDO 1999). Steife Gebilde setzen hingegen die Verformbarkeit herab und erlauben so eine Energieeinsparung, die aufgewandt werden müsste, sollten kontraktile Fasern unvorteilhafte Deformationen verhindern (GRASSHOFF 2014). Insbesondere bei den Tieren ist hierbei noch die Beweglichkeit der Lebewesen zu berücksichtigen. Bewegungen werden im hydraulischen System erzeugt, indem räumlich nahe beieinanderliegende kontraktile Fasern sich verkürzen und so das Volumen in ihrer Umgebung verringern. Die Flüssigkeit weicht aus und erweitert mit ihrem Druck an allen anderen Stellen das Volumen, wobei dort liegende Fasern verlängert werden (GRASSHOFF, GUTMANN & SCHÄFER 1994). Man spricht hier von antagonistischer Anordnung, die sowohl in der Zelle, als auch im Muskelsystem verwirklicht ist (GRASSHOFF 2014). Starre Elemente, die in die hydraulische Konstruktion eingebaut werden sind energetisch für die Bewegungstätigkeit eines Organismus günstig: ein Weichkörper ohne Versteifungen muss jede Muskelkontraktion durch eine andere Ausbalancieren, damit die Körperform erhalten bleibt und eine gerichtete Fortbewegung stattfinden kann. Starre Bereiche behalten ihre Form ohne Energieaufwand, sie ökonomisieren also den Betrieb (GRASSHOFF 2014).
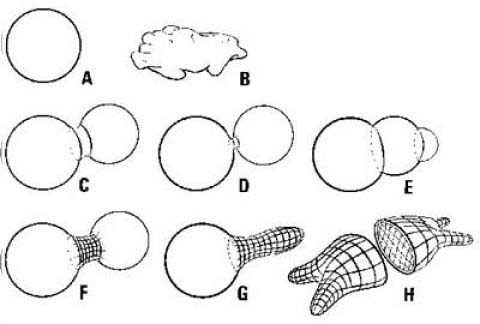
Abb.3: Formbildung bei Zellen.
A Von selbst stellt sich die Kugelform ein, B Unterfüllung hat Unbeweglichkeit zur Folge, C/D/E Teilung durch Schnürung, F/G teilweise Schnürungen lassen teilkugeln in verschiedener Form entstehen, H Eine aktive, phagozytierende Zelle muss über eine vollkommene räumliche Verspannung verfügen. Quelle: Gutmann (1995)
„Lebende Organismen sind als Konstruktionen hydraulische Apparate. Sie bestehen aus wässrigen Lösungen in flexiblen Membranen. Alle ihre Bewegungen sind aktive, von innen bewirkte Deformationen des Körpers. Diese werden durch den Einfluss von Muskeln, oder in einzelnen Zellen durch das Gleiten von Aktion- und Myosinfasern, hervorgebracht. Schon die Form der Lebewesen ist erzeugt, durch Einwirkung mechanischer Bauteile mechanisch erzwungen. Biologische Form ist nicht umsonst, sondern Ausdruck energiezehrender Leistung. Gegenstand der erklärenden Morphologie ist somit nicht die Form von Lebewesen, sondern das Gefüge der die Form und Bewegung bewirkenden Bauteile in ihrem Zusammenhang.“ (Gutmann 1987)
Interessant ist die Feststellung VOGELS (1991), dass dieser Umstand in den meisten Publikationen nicht beachtet wird. Wenn beispielsweise Hartteile am Körper, wie Gehäuse, gefunden werden, so werden die Antworten dafür immer in diversen Umweltbeziehungen gesucht, z. B. als Schutz vor Raubfeinden. Fast niemand fragt sich aber, „ob derartige Skelette nicht auch Bedeutung für den ‚Innenbetireb‘ des Organismus haben könnten, für ihre innere ‚Maschinerie’“ (S. 7).
Auch Einzeller und Bakterien haben vielfältig definierte Formen und sind hydraulische Gebilde, da die Zelle aus einer Membran und einer flüssigen Füllung bestehen. Für Bewegungen sorgt das Cytoskelett (BEREITER-HAHN, 1991, BEREITER-HAHN & STROHMEIER 1987, GUTMANN 1983). Ökonomisierungen können dabei durch die Einlagerung einer festen Zellwand z. B. bei Bakterien oder Pflanzenzellen entstehen (BENDEROTH, EDLINGER & GUTMANN 1995). BONIK (1978, 1979), der sich mit der aus überwiegend Siliciumdioxid bestehenden Hülle der Diatomeen (Kieselalgen) befasst hatte, konnte z. B. aufzeigen, dass ein Entfernen des Gehäuses dafür sorgt, dass sich die Kieselalgen abkugeln. Das Vorhandensein eines Gehäuses sorgte aber für eine Reduktion des Cytoskeletts.
Interessant ist auch, dass die Hydraulik der Körperkonstruktion in 300 Jahren Forschung kaum beachtet wurde. GUTMANN & BONIK (1980) gehen in ihrem Aufsatz „Borelli und die Folgen“ diesem Versäumnis genauer auf den Grund. Borelli hatte im 17. Jh. das Knochenskelett und die Muskeln des Menschen als zusammenwirkende Teile, also als funktionelles Ganzes erkannt. Eine Kontraktion des Muskels verstellt die Knochen über ein zwischen ihnen liegendes Gelenk. Die weitere Forschung blieb auf solche einfachen Zwei-Schritt-Ereignisse des Skelett-Muskelsystems beschränkt. Dabei vergaß man, dass Weichteile, seien es Darm und Herz, aber auch ein Wurmkörper auf eine andere Weise bewegen (GUTMANN & BONIK 1980, GRASSHOFF 2014).
„Es gibt in einer Hydraulik keine einfachen Antagonismen, kein simples Gegenspiel von zwei Muskelgruppen. Alles wirkt mit allem zusammen. Das kann man nicht sehen oder unmittelbar erfassen. (…) Man kann nicht aus den Systemteilen [wie im Fall von zwei Knochen, zwei Muskeln] addierend die Elemente zur Hydraulik verknüpfen. Hydraulik ist nicht ein addierter Verband, sondern ein geschlossenes Gesamtsystem“ (GUTMANN & BONIK 1980: 271)
BONIK, GRASSHOFF & GUTMANN (1976) geben noch einen weiteren Grund für das mangelnde Verständnis:
„Angesichts der Bildungssituation und der geistigen Traditionen des vorigen Jahrhunderts ist dies aber nicht verwunderlich: dem Gebildeten waren Technik und technisches Denken fremd; technisches Konstruieren war einem ‚Mittelbau‘ von Ingenieuren zugewiesen, der Gebildete der Universität sah in Technik kein geistiges Problem. Unter diesen Umständen verwundert es nicht, dass die inneren Mechanismen von Tieren (und Pflanzen) erst recht nicht erkannt wurde und damit kaum Fragestellung war.“ (BONIK, GRASSHOFF & GUTMANN 1976: 130)
Organismen als Energiewandler
Organismen werden als energiewandelnde Konstruktionen verstanden und versorgen sich durch ihre Aktionen selbst mit Energie. Organismen müssen ihre Lebensleistungen mittels Energieverbrauch erbringen. Sie nehmen Energie auf und müssen diese wieder nach außen abgeben. Dazwischen liegt die organismische Aktion, die Lebenstätigkeit, die durch Energiewandel hervorgebracht wird (Gutmann 1990, 1993, 1997, Gutmann & Bonik 1981). Dieses Prinzip, unter dem Fachbegriff des Bionomie-Kreislaufs zusammengefasst, wird in Abb. 4 dargestellt. Das aufgenommene Material wird im Metabolismus innerhalb der geschlossenen Konstruktion aufgeschlossen und in eine andere Energieform (ATP als Energieträger) umgewandelt. Diese Energie wird zur Selbstherstellung, Reparatur und zum Antrieb genutzt. Hier wird also auf Ebene der Biomechanik die chemische Energie in mechanische umgewandelt. Auf der dritten Ebene, der Hydrodynamik, erfolgt die geordnete Aktivität des Organismus durch die vom Bewegungsapparat vorgegebenen Restriktionen, die Investition eines Teils der Energie zur Reproduktion sowie den Erhalt und das Wachstum der bestehenden Struktur. Dieses Zusammenspiel bezeichnet man als Bionomie (BONIK & GUTMANN 1980, GUTMANN 1990, 1993, GUDO et al. 2006).
„Organismen stellen sich als bionome, d. h. sich selbst versorgende, selbstentsorgende und durch Abgliederung von Untereinheiten Reproduktion betreibende Maschinen dar“ (GUTMANN 1997)
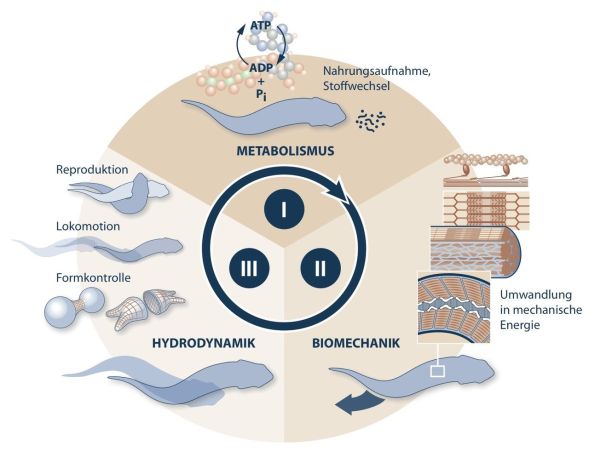
Abb. 4: Bionomiekreislauf der Organismen: I. Aufgenommene Nahrung wird nach chemischen Prinzipien aufgearbeitet und genutzt. II. Im Rahmen der Konstruktion erfolgt auf der biomechanischen Ebene Wandlung der chemischen und mechanische Energie, etwa durch das Gleiten der Aktin- und Myosin-Fasern in den Muskeln. Bewegung entsteht aus der Deformation der Konstruktion. III. Die Deformation muss in Antrieb und Verhalten umgesetzt werden, in dem koordinierte Organe (zum Beispiel das Nervensystem) steuernd eingreifen.
Auf jeder Ebene gelten eigene Prinzipien und Gesetzesmäßigkeiten.
Organismen nehmen also Nahrung aus der Umwelt aktiv auf, wandeln diese in verwertbare Energie um und betreiben damit die Konstruktion. Die aus der Nahrung gewonnene Energie wird z. B. in Form von Wärme und Bewegung wieder aktiv in die Umwelt abgegeben. Dabei hat Energie jedoch die Tendenz zu dissipieren, sich also in allen Teilschritten des Energiewandels gleichmäßig zu verteilen. Solche beliebige Verteilung wird nur durch die Struktur der Organismen verhindert, die die Energieflüsse leitet.
Ausgehend aus dieser Betrachtungsweise werden Organismen in der Konstruktionsmorphologie als thermodynamisch operational geschlossene Systeme definiert. Hier stehen sie im Gegensatz zu anderen Auffassungen, die Organismen als offene Systeme betrachten (Gutmann 1990, 1997, Gudo 2007). Dies erfordert eine kurze Erläuterung:
Ein System in der Thermodynamik bezeichnet man einen abgetrennten Bereich der durch physikalische Größen (Druck, Temperatur etc.) definiert ist. Ein System, das sowohl Materie als auch Energie mit seiner Umgebung austauschen kann, nennt man offen; ein System, das Energie aber keine Materie mit seiner Umgebung austauschen kann, wird als geschlossen bezeichnet; und ein System, das weder Energie noch Materie austauschen kann, wird isoliert oder abgeschlossen genannt.
Laut der Konstruktionsmorphologie sind Organismen operational geschlossen. Das heißt, bei Organismen handelt sich um autonome, selbst-organisierte Systeme, die zwar mit der Umwelt interagieren, aber gemäß ihrer Konstruktion. Ähnlich einem offenen System können sie also mit ihrer Umgebung Materie und Energie tauschen. Aber die Energie fließt nicht einfach durch sie hindurch, sondern wird in vielstufiger Weise gewandelt und in Bewegung, Wachstum, Fortpflanzung und erneute Energieversorgung investiert. Der Energiefluss durch den Organismus läuft nicht von allein, sondern durch eine aktive (energiezehrende) Leistung des Organismus: Das heißt ein Organismus muss sich Nahrung beschaffen, diese in seinen Stoffwechselprozessen umwandeln und damit seine Lebensfähigkeit erhalten. Das Beschaffen neuer Energie (und Nahrung) kostet dabei immer bereits Energie, die schon zuvor aufgenommen worden sein muss. Organismen sind insofern autonom als ihr Aufbau darüber entscheidet, was sie aus der Umgebung aufnehmen, verwerten und wo sie leben können. Um es an einem (vereinfachenden) Beispiel zu verdeutlichen: Unsere Konstruktion erlaubt es nicht die Energiequelle Cellulose, dem Hauptbestandteil des Pflanzengewebes, zu nutzen. Entsprechend unserer Konstruktion nehmen wir andere Energiequellen zu uns. Ganz anders bei den Wiederkäuern (Antilopen, Schafe, Hirsche, Rinder): Ihre Konstruktion (Magen mit mehreren Kammern und entsprechender Darmflora) erlaubt es Cellulose aufzuspalten und nutzbar zu machen (hier findet ein Energiewandel statt). Entsprechend entscheidet ihr Aufbau darüber, was sie aus der Umgebung aufnehmen, verwerten und wo sie leben können. Die Kennzeichnung von Organismen als offene Systeme ist laut der Konstruktionsmorphologie falsch, weil sich so keine Systemgrenzen benennen lassen und die Tätigkeit des Organismus von anderen Vorgängen nicht unterscheidbar wäre (EDLINGER et al. 1991).
Durch diese Tatsachen unterscheide sich ein Organismus von offenen Systemen, so die Konstruktionsmorphologie.
Ich bin kein Experte der Thermodynamik sowie der Wissenschaftsphilosophie und kann diesbezüglich keine Bewertung geben, ob die Bezeichnung eines operational geschlossenen System rechtens ist.
Doch zumindest erscheint die Erkenntnis, dass Organismen Energiewandler sind und ihre Konstruktion darüber entscheidet, welche Energiequelle sie nutzen können, durchaus plausibel. Damit unterscheiden sich Organismen auch qualitativ von unbelebter Materie, ein Unterschied, der in der Physik irrelevant ist.
Organismen als kohärente Systeme
Dieser Energiewandel findet aber bei Organismen auf Ebene der makromolekularen Ordnung statt. Die molekulare Arbeit setzt sich nur deswegen in Lebensleitung um, weil die gleitenden Strukturen in einem mechanisch-zusammenhängenden Gefüge wirken. Organismen sind dabei als kraftschlüssig organisierte Gebilde definiert. Man spricht von Kohärenz (GUTMANN 1986, 1987, PETERS, MOLLENHAUER & GUTMANN 1971).
„Der Organismus ist als funktionelles Ganzes definiert, ein lückenloses Gefüge, das nirgends unterbrochen ist. Kräfte werden also kontinuierlich und ohne Unterbrechung weitergeleitet. Man spricht von einem kraftschlüssigen Gefüge. Diese Gefüge sind aufrechtzuerhalten und im funktionell-strukturellen Kontext zu bestätigen. Bindegewebe, Zellen und Organe sind ständigen degenerativen Prozessen ausgesetzt. Sie werden also ständig abgebaut und aktiv vom Organismus neu aufgebaut. … Dieser Zusammenhang geht weit über das hinaus, was man von technischen Apparaten kennt und was mit dem technischen Terminus kraftschlüssig erfasst werden könnte. Wir nennen diesen speziellen Zusammenhang bei Organismen daher Kohärenz. Der Unterschied zum Kraftschluss einer Maschine besteht darin, dass deren Kraftschluss unterbrochen und wiederhergestellt werden kann… Die kohärente Organisation hingegen existiert kontinuierlich und von einer Generation zur nächsten, in Form abgegliederter funktionaler Untereinheiten. … Die Kohärenz beschreibt den strukturellen Zusammenhang des Ganzen im organismischen wie im prozessualen Sinne. Kohärenzverlust bedeutet eine irreversible Unterbrechung des Transformationsprozesses. Daraus folgt, dass Insuffizienzen organismischer Konstruktion nicht von der Umwelt aus, sondern ausgehend von ihrem mechanischen Gefüge erklärt werden müssen.“ (Gudo et al. 2006)
Ein Organismus ist daher ein vom kleinsten Teil bis zur vollständigen Konstruktion lückenloses, kraftschlüssiges Gefüge. Dieses Gefüge wird immer erhalten und auch an die Nachkommen weitergegeben (Gutmann 1987, 1993, 1995, 1997).
„Die Nutzung der Energie bei gerichteter Aktion setzt neben Energiewandel, durch die Struktur der makromolekularen Ebene bestimmt, Einrichtungen voraus, die die Nutzung der freigesetzten Energie kanalisieren, also beliebig ungerichtete Kraftauswirkung unterbinden. Die Kanalisierung wird durch interne restringierende Strukturen bewirkt, die in strenger Beziehung zu den Kraftschluss sicherenden Einrichtungen bestehen. Die aus chemischer Energie gewonnene mechanische Aktion muss von der Ebene der Makromoleküle auf das arbeitende Gefüge übertragen werden, das sich ausformt und bei Aktionen deformiert. Diese Wirkung ist nur möglich, wenn alle Strukturen im operational geschlossenen Rahmen eingebaut sind und mechanische Kohärenz gewährleistet ist.“ (Gutmann 1997)
Das Beispiel der Kohärenz wird in Abb.5 verdeutlicht. Dabei werden Kräfte über Muskeleinheiten, Bindegewebe und grobe morphologische Strukturen auf das arbeitende Gefüge übergeleitet. So sind die kleinsten Einheiten der Muskulatur, die Sarkomere, die aus Aktin- und Myosinfasern bestehen, zu größeren funktionalen Einheiten, den Muskeln, zusammengeschaltet. Die Muskeln selbst sind in den Bindegewebsverband des Organismus eingelassene Kompartimente. Zwar wird die Muskelverkürzung durch Energiewandel bewirkt, aber die Muskelarbeit als solche besteht nicht nur aus deren Verkürzung. Eine sinnvolle Muskelaktion muss wiederholbar sein und dies ist nur möglich, wenn der Muskel wieder gedehnt werden kann. Diese Dehnung wird aber nicht aus der Molekularstruktur des verkürzten Muskel bewirkt. Erst durch die Integration in einen mechanischen Verband wird dafür gesorgt, dass sich der Muskel durch andere Muskeln, die nicht an der Bewegung beteiligt sind, dehnen kann. Es entsteht also ein Muskel-Antagonismus und nur aus diesem Gesamtzusammenhang lassen sich Bewegung und Muskelarbeit sinnvoll erklären. Aus der molekularen Struktur des Muskels alleine lässt sich Muskelarbeit nicht sinnvoll erklären (GUTMANN 1986, BONIK & GUTMANN 1980). Dieses Verständnis der Kohärenz hat wichtige Konsequenzen für die reduktionistisch ausgelegte Molekularbiologie.
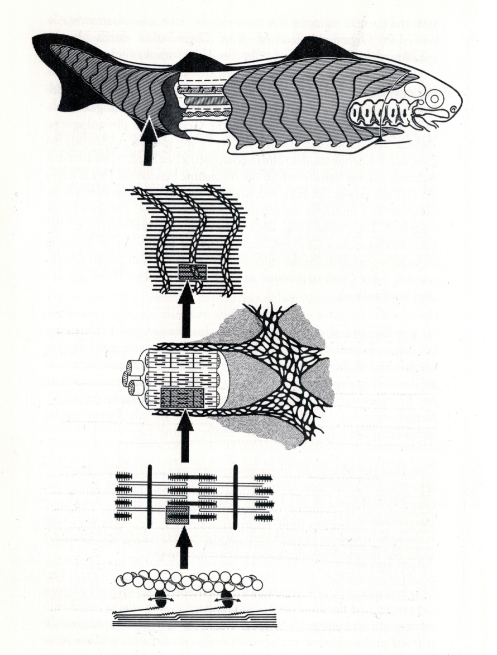
Abb. 5: Der Organismus als kohärentes Gefüge. Die energetische Powerung eines beweglichen Organismus durch chemo-mechanischen Energiewandel erfolgt auf der molekularen Ebene durch das ATP-verbrauchende Gleiten von Aktin-Myosin-Fasern aneinander. Mechanisch-motorische Leistung des Gesamtorganismus kommt durch molekulare energetische Powerung in der Weise zustande, dass die Energiewandlung im kohärenten Verband geschieht, der die Energie kanalisiert und in morphologische Strukturen auf den Gesamtkörper hochleitet. Quelle: Gutmann (1995)
Spezialisierung, Ökonomisierung und Differenzierung
Für die Evolution der Lebewesen sowie bei der Rekonstruktion einer Abwandlungsreihe gelten folgende Prinzipien: Spezialisierung, Ökonomisierung und Differenzierung (Grasshoff 2014).
Jedem Organismus ist nur eine begrenzte Energiemenge zur Verfügung gestellt. Diese Menge wird aber nicht durch die Ressourcen bestimmt, sondern durch die Konstruktion des Organismus selbst. Das Konstruktionsgefüge des Organismus bestimmt, wie viel Energie er aufnehmen und umwandeln kann. So befinden sich Organismen in einem ständigen thermodynamischen Ungleichgewicht. Dieser Zustand kann nur gewährleistet werden, dass sich der Organismus unter Energiewandlung ständig neue Nahrung (also Energie) zuführt. Der ständige spontane Abbau und das beständige Wirken des Organismus gegen diesen Abbau durch Materialaufnahme, führt zu ständig neuen Entropiezuständen, zu ständiger Veränderung des Körpers, zu kontinuierlichem Wandel und damit zur Evolution (Gutmann 1989, 1995, 1997). Bei der Ökonomisierung werden Stoffwechselvorgänge selbst ökonomisiert oder es werden entbehrliche Strukturen reduziert (z. B. Reduktion der Ringmuskulatur während der Ausbildung der Chorda zur Längenkonstanz). Unter einer Optimierung bzw. Spezialisierung ist eine Verbesserung hinsichtlich der Leistung zu verstehen (Grasshoff 2014). Während sich die Ökonomisierung auf das Konstruktionsgefüge bezieht, bezieht sich die Optimierung auf das Wirkungsgefüge der Organismen. Differenzierung ist hingegen gleichzusetzen mit strukturellen Veränderungen, die Teilen der Konstruktion neue Funktionen erlauben und so das Erschließen neuer Lebensräume erlauben. Differenzierung ist der Prozess, der neue Konstruktionen hervorbringt (Gudo 1999). Dabei bestimmt die Ausgangskonstruktion die Weiterentwicklung. Da Lebewesen nicht wegen Umbau geschlossen werden können, müssen Funktionsfähigkeit und Fortpflanzungsmöglichkeiten bei jeder Veränderung gewahrt bleiben. Evolution kann also nur durch kleinschrittigen Wandel erfolgen. Große Abwandlungen sind meist Missbildungen und der Organismus geht dann auf Grund seiner Konstruktion zugrunde. Ob ein Organismus in seiner Umwelt überleben und sich reproduzieren kann, hängt also von seiner Konstruktion ab (Gutmann 1989, 1995,1997a Gudo 1999, 2007). Es werden nämlich diejenigen Nachkommen selektiert, die nicht in der vorliegenden Umwelt zurechtkommen.
„Weil Evolution graduell verläuft, können neue Strukturen nicht spontan (aus dem Nichts) hinzukommen; es gilt: aus Nichts entsteht Nichts. Auch wenn in der Evolution scheinbar immer wieder neue Strukturen entstanden sind, so sind diese kontinuierlich entstanden, indem sich bereits vorhandene (wenig oder undifferenzierte) Gewebe zunehmend differenzierte und in neue Strukturen gebildet haben. Um die Grenzen des Wandels auszuloten, ist die Einbeziehung der Histologie (= Gewebekunde) und damit ein Blick auf die mikroskopische Organisation des Körpers unerlässlich. Wir wissen, dass undifferenzierte Gewebe sich zu vielfältigen Organen und Strukturen differenzieren können. Bereits differenzierte Gewebe aber haben ein deutlich eingeschränkteres Entwicklungspotential. … Übertragen auf die Evolution bedeutet dies, dass zwar Flossensäume zu Flossen und diese zu Beinen werden können, wenn aber Beine einmal ganz abgebaut sind, gibt es auch keine organische Grundlage mehr, aus denen Extremitäten neu entstehen könnten.“ (GUDO 2007: 30).
Evolutionsabläufe sind somit Abfolgen von Konstruktionen und für alle Schritte wird die Unumkerhbarkeit explizit begründet (GRASSHOFF 1994a).
Die Konsequenzen der Konstruktionsmorphologie haben aber auch Folgen für GOULDS (1999) Kontingenztheorie, die davon ausgeht, dass sich Evolution nicht wiederholen würde und völlig neue Organismen entstehen würden. Tatsächlich würden nicht identische Organismen entstehen, wahrscheinlich auch nicht dieselben Radiationen einzelner Gruppen. Jedoch würden ähnliche Ergebnisse zu erwarten sein und bei einem Neu-Ablauf der Evolution ähnliche Konstruktionen entstehen (GUTMANN 1996a,b, GUDO 2008). Evolution hat damit kein Ziel, wohl aber auf Grund der Konstruktionsbedingungen der Lebewesen eine Richtung.
„Es lässt sich – unter Rückgriff auf die Erkenntnisse der FET [=Frankfurter Evolutionstheorie = Konstruktionsmorphologie] – der Rahmen geben, innerhalb dessen die Evolution weiterlaufen wird. Wenn die Prinzipien bekannt sind, nach denen evolutionärer Wandel abläuft und rekonstruierbar ist, so lässt sich auf einzelne Evolutionslinien über den weiteren Verlauf des Wandels eine näherungsweise Aussage treffen.“ (GUDO 2007: 32)
Die Möglichkeiten des evolutionären Wandels wird somit von der Körperkonstruktion sowohl eröffnet als auch begrenzt.
Organismus und Molekularbiologie
Dieser Ansatz betrachtet also Organismen als Ganzes, da sie mehr darstellen als die Summe ihrer Einzelteile oder als Merkmalsträger. Entsprechend lässt sich ein Organismus nicht erklären, wenn man ihn in seine Einzelteile zerlegt. So wie sich die Funktion eines Ottomotors nicht aus der chemischen Verbrennung des Benzins erklären lässt, lässt sich ein Organismus auch nicht aus seinen molekularen Strukturen, z. B. seiner Gene, erklären. Organismen können so auch nicht auf „Gen-Vehikel“ reduziert werden (vgl. DAWKINS 1976). Da lebende Organismen in den Dimensionen des Raumes und der Zeit existieren, lassen sie sich nicht aus der eindimensionalen Struktur der DNA ablesen (Rose 2000, VOGEL 1991). Dies zeigt alleine schon das oben erwähnte Beispiel der Muskelkontraktion durch Muskelantagonisten. Aus der Aktin- und Mysionstruktur (bzw. der dafür kodierenden Gene) lässt sich die Muskelarbeit nicht sinnvoll beschreiben, erst im Verband ist sie erklärbar. Wir haben es mit den Organismen um einen höheren und komplexeren Verband der Materie (vgl. ROSE 200: 109 – 111) zu tun, bei der neue Interaktionen und Beziehungen entstehen, die man nicht reduktionistisch durch Zerlegung in ihre Eigenbestandteile erklären kann. Folgerichtig kritisiert die Konstruktionsmorphologie den Reduktionismus der Molekularbiologen. Die Bedeutung der Molekularbiologie wird nicht ignoriert, ihr reduktionistischer Monopolanspruch zur Erklärung von Form und Struktur wird in der Frankfurter Evolutionstheorie jedoch in Frage gestellt.
Gene enthalten zwar wichtige Materialinformationen, aber noch lange keine vollständige Bauinformation. Im genetischen Apparat und dessen Kodierung liegen nicht sämtliche notwendigen „Informationen“ für den Organismus vor (Gutmann 1997). So gibt es eine Vielzahl kleinerer und größerer Moleküle, wie Ionen, Polysaccharide und Proteine, ohne die die DNA nicht funktionsfähig wäre. Die Metapher der DNA als Buch des Lebens entspricht nicht der Realität. DNA ohne zelluläre Umgebung ist nutzlos.
„Wir haben oft sagen hören, dass die Gene die ‚Informationen‘ enthalten, die ein Lebewesen spezifiziert. Dies ist aus zwei fundamentalen Gründen falsch: Erstens wird dabei das Vererbungsphänomen verwechselt mit dem Replikationsmechanismen gewisser Zellbestandteile (DNS) mit großer transgenerationaler Stabilität. Es ist zweitens falsch, weil wir mit der Behauptung, dass die DNS das Notwendige enthalte, um ein Lebewesen zu spezifizieren, die DNS (eine Komponente des autopoietischen Netzwerks) aus ihrer Einbindung in den Rest des Netzwerks herauslösen. Es ist jedoch dieses Netzwerk von Interaktionen in ihrer Gesamtheit, welches die Charakteristika einer bestimmten Zelle konstituiert und spezifiziert – und nicht etwa nur ein Bestandteil dieser Zelle. Dass Modifikationen in diesen Komponenten, den Genen, dramatische Konsequenzen für die Struktur einer Zelle haben, ist wohl wahr. Der Fehler in der oben ausgeführten Argumentation besteht darin, wesentliche Beteiligung mit alleiniger Verantwortung zu verwechseln.“ (MATURANA & VARELA 1987: 78)
Molekulare Systeme powern den Organismus zwar mit Material und Energie, jede Form ist letztendlich mechanisch bedingt:
„Die Wirkung der Gene auf die Morphologie geschieht sicher über biochemische Verbindungsglieder, aber diese wirken auf einen Verband, der eigenen Gesetzen, denjenigen der Hydromechanik, gehorcht. Das bedeutet, dass die biochemische Wirkkette der Genwirkkette notwendige Bedingungen liefert, dass das Resultat von den organisatorischen Prinzipien der morphologisch-mechanischen Ebene abhängt. Variabilität ist also nicht einfach eine Umstellung der makromolekularen Mechanismen, sondern die Transformation eines mechanischen Gefüges.“ (Gutmann 1995)
Die Bedeutung der Molekularbiologie wird nicht ignoriert, ihr reduktionistischer Monopolanspruch zur Erklärung von Form und Struktur wird in der Frankfurter Evolutionstheorie jedoch in Frage gestellt.
„Struktur und Organisation ohne molekulare Mechanismen sind tot, können mangels Material- und Energiezulieferung nicht arbeiten, sind starr zur Reparatur und zur Reproduktion unfähig. Molekulare Mechanismen sind für alle Lebensabläufe absolut indispensabel, aber sie konstituieren nicht die Struktur und den organisatorischen Zusammenhang; das bedeutet, dass man nicht durch immer weitergetriebene molekulare Analyse und die Aggregation partikulärer Abläufe und Mechanismen die Einsicht in Struktur und Organisation sowie ihre Prinzipien erreichen kann.“ (Gutmann1995b)
Organisation und Struktur haben ihre eigenen Prinzipien: Hydraulik, operationale Geschlossenheit, Energiewandel, Kohärenz etc., die nicht durch die Molekularbiologie erklärt werden können. Letztere dient lediglich zur Material- und Energielieferung. Die Vorstellung, dass Gene direkt für komplexere Strukturen kodieren, ist eine Fehlkonzeption (LANGE 2012). Dieser Vorstellung schließt sich auch WEST-EBERHARD (2003) an, die in ihrem Werk nachweist, dass genotypische Veränderungen den phänotypischen folgen. Die Embryonalentwicklung wird als Motor der Evolution gesehen und die Embryonalentwicklung ist das Ergebnis vieler epigenetischer Regulationsprozesse. Änderungen dieser Prozesse werden durch Umwelteinflüsse angestoßen. Auch wenn WEST-EBERHARD und LANGE nicht konstruktionsmorphologisch argumentieren, da auch die Embryonalentwicklung konstruktionsmorphologisch analysiert werden muss und nicht als verkürzter Ablauf der Phylogenese wie es HÄCKEL verstand (vgl. BONIK, GRASSHOFF, GUTMANN 1978, 1979a, b, 1981), sind ihre Aussagen Folge der Konstruktion der Organismen. Da das Konstruktionsgefüge der Organismus bestimmt wie viel Energie er aufnehmen und umwandeln kann, befinden sie sich in einem ständigen thermodynamischen Ungleichgewicht. Ihr Zustand kann nur gewährleistet werden, dass sich der Organismus unter Energiewandlung ständig neue Energie zuführt. Der ständige spontane Abbau und das beständige Wirken des Organismus gegen diesen Abbau durch Materialaufnahme, führt zu ständig neuen Entropiezuständen, zu ständiger Veränderung des Körpers, zu kontinuierlichem Wandel und damit zur Evolution (GUTMANN 1989, 1995, 1997, GUDO 2007). Die Kopierfehler bei der Vererbung sind also zwangsläufige Folge der physikochemischen Eigenschaften der reagierenden Verbindungen (GRASSHOFF 1993a).
Hier ist die Ursache jeglicher Variation (ob genetisch oder epigenetisch) zu suchen. Sie haben eine thermodynamische Grundlage. Damit ist Variation nicht folge zufälliger Mutationen, sondern bedingt durch die Konstruktion der Organismen. Jede Abwandlung des Gefüges muss dabei die konstruktiven Mechanismen des Energiewandels intakt lassen und kann nur nach Maßgabe der Körperkonstruktion geschehen. Damit wird die Richtung der Umwandlung durch den Aufbau des Organismus bestimmt (EDLINGER, GUTMANN & WEINGARTEN 1989). Dies gilt sowohl für adulte Organismen als auch für Embryonen. In dieser Hinsicht muss auch das von WEISMANN (1892a,b) aufgestellte Konzept der Keimbahn neu interpretiert werden. WEISMANN konnte nachweisen, dass sich in der Embryonalentwicklung vieler Tiergruppen die Urkeimzellen früh absondern und anders als die somatischen Zellen nicht an der Entwicklung des Organismus beteiligt waren. Entsprechend bestehen vielzellige Organismen aus den Keimzellen, die für die Fortpflanzung nötig sind und den somatischen Zellen, die die Körperfunktionen erfüllen. Die Zellenabfolge aus der die Keimzellen hervorgehen bezeichnet man dabei als Keimbahn. Die von der Keimbahn abzweigenden somatischen Zellen bilden den Körper (das Soma). Da durch die Keimbahn eine ununterbrochene Folge von Keimzellen entstehen, gelten diese als potentiell unsterblich. Die Körperzellen unterliegen dagegen einem natürlichen Tod (GRIESEMER 2002). Eine wichtige Folge dieser Beobachtung ist, dass die Keimzellen von den Somazellen nicht beeinflusst werden, womit die Vererbung erworbener Eigenschaften als widerlegt galt. WEISMANNS Vererbungstheorie bildet die theoretische Grundlage des „Zentralen Dogmas der Molekularbiologie“ (CRICK 1970). Grundaussage ist, dass keine Information von Protein zu Protein oder zu Nukleinsäure möglich ist. WATSON (1965) gibt eine vereinfachte Version des Zentralen Dogmas wider (DNA -> RNA -> Protein), während CRICK auch die Möglichkeit RNA -> DNA (z. B. beim Enzym Reverse Transkriptase) und DNA -> Protein zuließ. So richtig die Beobachtung von WEISMANN ist, so ist sie zum einen nicht universell, da Pflanzen, Pilze und einige Tiergruppen sowie die Einzeller keine Keimbahn entwickeln. Zum anderen, wenn man von einem kohärenten und operational geschlossenen energiewandelnden Organismus ausgeht, ist eine Unabhängigkeit der Keimzellen vom Rest des Organismus nicht möglich. Auch die Keimzellen sind in das kohärente System integriert, nicht zuletzt weil auch Keimzellen mit Energie gepowert werden müssen. Morphologisch lässt sich dies daran zeigen, dass jede Eizelle von Nährzellen (Keimbahn-Ursprung) und Follikelzellen (somatischer Ursprung) umgeben ist (WOLPERT 2007). Diese Zellen versorgen die Eizelle mit RNAs, Proteinen, Nährstoffen, Neurotransmittern und Hormonen. Diese Stoffe sind für die spätere Embryonalentwicklung unerlässlich. Die Konzentration dieser Stoffe, sowie deren Transport über die Nähr- und Follikelzellen wird über das neuro-hormonelle Netzwerk des Organismus gesteuert (CABEJ 2012, 2013). Nährstoffe wie Dotterproteine werden bei Vögeln und Amphibien in den Leberzellen gebildet und gelangen über das Blut zu den Eierstöcken. Dort werden die Proteine von der Eizelle durch Phagozytose aufgenommen (WOLPERT 2007). Von CRICK wird diese Einbahnstraße des Informationsflusses reduktionistisch verstanden, da die DNA zum Mastermolekül erhoben wird (ROSE 2000). Die Information die aber in den Nukleinsäuren gespeichert ist nur ein Teil der für die Proteine notwendigen Informationen. Die einzige Information, welche sich in der DNA befindet ist die Anordnung der Aminosäuren in einem Protein sowie entsprechende Regulationssequenzen. Aus der Anordnung der Aminosäuren alleine ergibt sich jedoch kein Bauplan eines Organismus (GUTMANN & BONIK 1981). Dies erkennt man alleine daran, dass ein Protein nur dann seine Wirkung entfalten kann, wenn es gefaltet ist, also eine dreidimensionale Struktur aufweist. Die Faltung der Proteine ist aber zu einem wesentlichen Bestandteil abhängig vom zellulären Umfeld (LEWONTIN 2002, vgl. Abb. 6).
Auch die Genregulation, also wann ein bestimmtes Gen aktiviert oder abgeschaltet wird, ist in der DNA nicht codiert. JABLONKA & LAMB (2004) konnten auch zeigen, dass für die Vererbung „erworbener Eigenschaften“ eine Rückübersetzung von Protein zu DNA nicht notwendig ist, da die „erworbenen Eigenschaften“ keine Veränderungen der Aminosäurensequenz erfordern. Bei der Reaktion der Organismen auf veränderte Umweltbedingungen ändert sich nicht die Aminosäurensequenz, sondern die Menge bestimmter Proteine durch Genregulation. Genregulation ist ein komplexes Wechselspiel zwischen Genen, Organismus und Umwelt (vgl. CABEJ 2012, 2013, JABLONKA & LAMB 2004, GILBERT & EPEL 2009, LANGE 2012, LEWONTIN 2002). Desweiteren konnte gezeigt werden, dass bestimmte Proteine, sogenannte Prionen, in der Lage sind andere Proteine mit ähnlicher oder gleicher Aminosäurenzusammensetzung in ein Ebenbild ihrer eigenen Struktur umzubilden (JABLONKA & LAMB 2004, HALFMANN & LINDQUIST 2010). Dadurch wird der Informationsfluss von den Nukleinsäuren (DNA, RNA) zu den Proteinen weder falsch noch unnötig. Doch für die Konstruktion des Organismus ist dieser nicht entscheidend, da weder in den Nukleinsäuren noch in den Proteinen die Information für die Konstruktion der Organismen codiert ist.
„Die Prinzipien der Körpermechanik bestimmen was an molekularer Organisation möglich ist“ (GRASSHOFF 1993a).
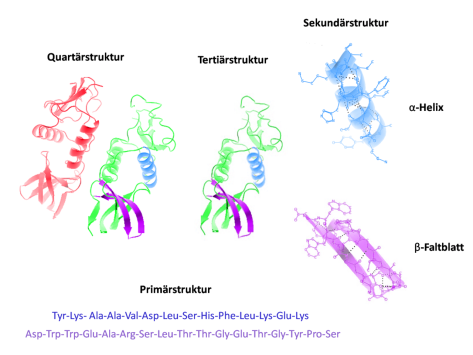
Abb. 6: Die unterschiedlichen Strukturen eines Proteins. Die DNA enthält nur die Information für die Primärstruktur der Proteine, also die Reihenfolge der Aminosäuren (der Bausteine der Proteine). Durch die Wechselwirkungen der Wasserstoffatome in den Aminosäuren entstehen die Sekundärstrukturen, die sich wiederrum in Tertiär-Strukturen und Quartärstrukturen verbinden. Diese Strukturen sind u. a. nicht nur abhängig von der Reihenfolge der Aminosäuren, sondern z. B. auch vom Milieu des Zellplasmas (pH-Wert, Bewegung des Zellplasmas, Enzyme etc.). Wenn die DNA also noch nicht einmal die Informationen für die (funktionelle) Struktur eines Proteins enthält, dann erst recht nicht für den Bauplan eines Organismus.
Es ist tatsächlich eher so, dass molekulare Mechanismen strukturabhängig verlaufen. Theoretisch gesehen könnte jedes Bauteil aus einem anderen Material hergestellt werden:
„Jedes Bauteil könnte auch aus einem anderen Material hergestellt werden. Man kann zudem mit dem gleichen Betriebsstoff unterschiedliche Konstruktionen treiben. Benzin lässt sich in Viertaktern, in Zweitaktmotoren, aber auch in Turbinen und im Raketentrieb nutzen. Unterschiedliche Materialien können den meisten Konstruktionsbedingten Anforderungen gerecht werden. Auch vom benutzten Betriebsstoff kann nicht auf die Maschinerie geschlossen werden. Wie ein Betriebsstoff wirkt und wie seine Energie umgesetzt wird oder werden kann, hängt immer vom angetriebenen Gefüge ab, das so gestaltet sein muss, dass die Energie aus dem Betriebsstoff freigesetzt werden kann und die Kräfte in der Konstruktion so weitergeleitet werden, dass sich sinnvolle Aktion ergibt. Welche Energieform sich auf den verschiedenen Stufen der arbeitenden Konstruktion zeigt, welche Leistung letztlich bewirkt wird, das ergibt sich aus dem Resultat der morphologischen Organisation, also aus dem Aufbau und dem Wirkungsgefüge der Maschinerie.“ (Gutmann 1995b: 63)
Als Beispiel kann das Molekül ATP (Adenosintriphosphat, Abb. 7) genommen werden, dessen Spaltung in ADP + P (Adenosindiphosphat + Phosphat) Energie freisetzt, um biologische Aktion zu betreiben. ATP gilt dabei als universeller Energieträger, welcher in Stoffwechselprozessen gebildet und vielfältig eingesetzt wird. Die Nutzung der Energie, welche durch die Spaltung von ATP hervorgebracht wird, ist jedoch strukturabhängig:
„Wie die Energie genutzt wird, entscheidet weder die ‚energiereiche‘ Verbindung selbst, noch sind die Mechanismen, die für die Synthese verantwortlich sind, für das Nutzungsgeschehen bestimmend. Der Wirkort in seiner Struktur und das ihn umgreifende Gefüge samt der supramolekularen Organisation determinieren die Wirkung der Energie und legen die Art der biologischen Leistung fest. Das gleiche ATP wird hydraulische Wirkungen im Cytoskelett in Gang setzen können, Bewegungen durch Deformationen hervorrufen, einen Spindelapparat antreiben, der Chromosomen bei der Zellteilung verschiebt, glatte, schräggestreifte oder quergestreifte Muskeln zur Kontraktion befähigen. Die Wirkung dieser Muskeln, die nur bei Energiezufuhr zu arbeiten vermögen, hängt vom konstruktiven Verband, seiner Organisation mitsamt den durch ihn institutionalisierten Antagonismusbeziehungen ab. Über keinen molekularen Prozess ist das Vorliegen von antagonistischer Muskelwirkung im Verband zu begründen, oder die im Bauplan realisierte antagonistische Beziehung abzuleiten.“ (Gutmann 1995b: 66)
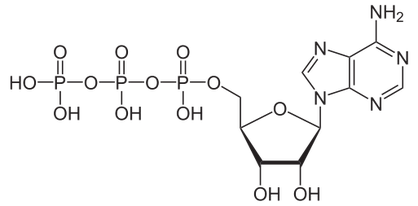
Abb. 7: ATP (Adenosid-tri-Phosphat), der universelle Energieträger bei Organismen. Zu welchem Zweck dieser Energieträger genutzt wird, hängt von der Struktur ab. Quelle: wikipedia
Komplexe Metabolismus- und Syntheseprozesse sind an Struktur und topologische Geschlossenheit gebunden (HAROLD 1986, 1990). INGBER (1993a, b) und RESNIK et al. (1993) konnten z. B. zeigen, dass DNA von der mechanischen Struktur her beeinflusst werden. Morphologisch gleiche Strukturen können auch durch verschiedene molekulare Signalwege und Regulationsmechanismen generieren, wie die Entstehung des Eiaplageapparates der Nematoden Caenorhabditis elegans und Pristionchus pacificus zeigen (STERBERG 2005, Tian et al. 2008, Zheng et al. 2005). Diese Tatsache, bestätigt zum einen GUTMANNS obige Aussage, dass jedes Bauteil von einem anderen Material geschaffen werden kann. Zum anderen zeigt sich auch hier, dass molekulare Prozesse den konstruktionsmorphologischen untergeordnet sind. Eine besondere Strukturabhängigkeit molekularer Prozesse liefert auch das Cytoskelett. BEREITER-HAHN (1994) konnte z. B. auch nachweisen, dass Enzyme der Glykolyse an das Cytoskelett gebunden sind. Das Cytokelett spielt auch eine Rolle bei der Regulation der DNA-Replikation (CASAS-DELUCCHI & CARDOSO 2011, THYBERG 1984, CABEJ 2013), der Genexpression (ROSETTE & CARIN 1995, HESKETH & PRYME 1991, OLSON & NORDHEIM 2010; CABEJ 2013) und der Zellteilung (CABEJ 2013). Auch der Transport und die Position maternaler RNAs und Proteine in der Eizelle wird über das Cytoskelett reguliert (CABEJ 2012, 2013).
„Eine vollständige Garnitur von DNS lässt auch bei Zugabe von allen nötigen chemischen Stoffen niemals einen Organismus entstehen. Bei einer normalen Entwicklung eines Lebewesens liegt nie nur die sogenannte Erbinformation vor, sondern immer eine ganze lebende Zelle. In der Zelle und ihrem Stoffwechsel sorgen Regulationsvorgänge dafür, dass die ‚richtigen‘ Teile des Erbapparates angeschaltet werden und ihren Einfluss ausüben. Die Erbmechanismen und deren Steuerung sind also nicht absolut vorgeordnet, sondern in das Geschehen der lebenden Zelle und in ihre Funktionskreise einbezogen. Das Erbgut ist nicht ein unabhängiges Meisterprogramm oder eine Kommandostelle für den Aufbau, es ist selbst sehr abhängig.“ (Gutmann & Bonik 1981: 123)
Die Prozesse der Molekularbiologie und die Entstehung erblicher Varianzen wird in einem späteren Kapitel vorgestellt. Es wurde aber schon hier dargestellt, dass Molekularbiologie alleine nicht ausreicht, um Evolution zu verstehen.
Konstruktionsmorphologie und klassische Morphologie
Die Erbsubstanz, wie alle anderen molekularen Mechanismen sind also in das kohärente Gefüge des Organismus mit eingebaut. Wenn man diesen Umstand mit einbezieht, entsteht ein völlig neuer Organismusbegriff. Organismen werden hier als funktionierende Körperkonstruktionen betrachtet und nicht als Merkmalsträger, wie dies in der traditionellen Stammesgeschichtsforschung (Kladistik) üblich ist. Hier haben sich zunehmend Widersprüche zur idealistischen Morphologie und zur phylogenetischen Systematik ergeben. Wir haben zu Beginn des Artikels dargestellt, dass die phylogenetische Systematik bzw. Kladistik durch ihre Unterscheidung in apomorphe und plesiomorphe versucht die Verwandtschaftsbeziehungen der Organismen zu klären und so Evolutionsabläufe zu rekonstruieren.
Das erfordert jedoch zum einen genaueste Kenntnis über die Morphologie der zu beschreibenden Gruppen. Zum anderen besteht jedoch die Gefahr, dass man sich vor lauter Merkmalen nicht so recht entscheiden kann, was ein gutes oder schlechtes Merkmal ist. Hinzu kommt, dass der Organismus in beliebig viele Merkmale aufgelöst werden und so als Forschungsobjekt der Biologie erheblich undefiniert und für bestimmte Fragestellungen sogar zerstört werden kann. Die synthetische Evolutionstheorie hatte nie einen funktionellen Organismusbegriff, sondern reduziert diesen auf Gene und ihre Wirkung. Die Körperkonstruktion und ihre Funktion (sowie die Entwicklungsbiologie) werden ignoriert oder als blackbox gesehen. Organismen werden auf die Summe ihrer Anpassungsmerkmale reduziert, die durch die natürliche Selektion geformt werden (GRASSHOFF 2014). Die traditionelle Morphologie, von der Konstruktionsmorphologie als Gestaltmorphologie genannt, sieht die Form als gegeben an. Dabei werden Form und Funktion auseinander gehalten. Die Form wird als das primäre, die Funktion als das sekundäre betrachtet. Die Ebene der Körpermechanik wird dabei weder wahrgenommen noch erwartet. Der Begriff der Hydraulik und das Hydroskelett wird allerhöchstens als Spezialfall, z. B. beim Regenwurm, erwähnt. (GRASSHOFF 1994a). Für Evolutionsreihen ist in der Gestaltmorphologie ein Additions-Denken typisch. Für die Rekonstruktion des Evolutionsverlaufes werden Gestaltreihen entworfen, bei der komplexere Formen phylogenetisch jünger sind, als einfachere Formen (GRASSHOFF 1994a). Es werden also neue Merkmale hinzuaddiert ohne zu bemerken, dass die Struktur der Organismen durch die Konstruktionsprinzipien der hydraulischen Organisation bestimmt sind (GRASSHOFF 1994a). Jedoch kann der Organismus in beliebig viele Merkmale aufgelöst werden und so als Forschungsobjekt der Biologie erheblich undefiniert und für bestimmte Fragestellungen sogar zerstört werden. Diese Methodik findet in der Homologieforschung und der daraus entstehenden phylogenetischen Systematik (Kladistik) ihren Höhepunkt.
„Hennigs Theorem (…), beruht im wesentlichen auf den (sonst nicht näher) definierten Begriff des Merkmals. Die Konsequenz daraus ist aber (…), dass nicht alle vorhandenen Merkmale für die Hennigsche Systematik benutzbar sind. Die Frage ist nämlich, wie plesiomorphe bzw. apomorphe Merkmale erkannt werden können, wie also die Merkmalswertung erfolgt.“ (Bonik 1981)
War die idealistische Morphologie zu ihrer Zeit noch ein wissenschaftlicher Fortschritt, der eine Ordnung in das Chaos der Organismen brachte, so wurde mit zunehmenden Erkenntnisgewinn über den Bauplan der Organismen, allen voran der Tiere, dieser in immer mehr Merkmale aufgelöst bis er als Forschungsgegenstand völlig verschwand. Der Organismus wurde nicht mehr als Ganzes gesehen sondern als Träger plesiomorpher (ursprünglicher) und apomorpher (neuer, abgeleiteter) Merkmale, wie ihn die Kladistik beschreibt (vgl. HENNIG 1950, 1966). Im deutschsprachigen Raum war es vor allem REMANE (1952) der die Homologiekritierien einführte und zur Lehrbuchmeinung wurden. Da der Organismus jedoch in beliebig viele Merkmale aufgelöst werden kann, ist es oft schwer zu ermitteln welches der Merkmale ursprünglich oder abgeleitet ist, bzw. ob das Merkmal auf einen gemeinsamen Vorfahren zurückgeht (also eine Homologie ist) oder nicht. Die Kladistik bringt die festgestellten Merkmale in ein formales Schema. Diese Arbeitsweise eignet sich für die Systematik, jedoch nicht für eine zeitliche Abfolge des Evolutionsverlaufs. Die Konstruktionsmorphologie negiert auch nicht den Nutzen der Kladistik für die Systematik. Es macht keinen Sinn die Taxonomie völlig abzuschaffen, da sie für die Einteilung der Lebewesen notwendig. Jedoch wird der absolute Anspruch mit der ordnenden Systematik, fußend auf der idealistischen Gestaltmorphologie, Evolutionslinien zu deuten abgestritten (vgl. GRASSHOFF 1993a, 1994a). Dies liegt u. a. daran, dass weder die HENNIGSCHE Systematik, noch die REMANES entscheidende Kriterien dafür liefern, wie die Lesrichtung der Merkmalsreihungen zu bestimmen ist (vgl. FRANZEN et al. 1973, PETERS & GUTMANN 1971, GRASSHOFF 1994a). Hinzu kommt, dass die Unterscheidung zwischen Plesiomorphie und Apomorphie um Wahrscheinlichkeitsentscheidungen handelt. Desweiteren werden keine funktionellen Interpretationen der Befunde verlangt (PETERS & GUTMANN 1971). Auch HASZPRUNAR (1994: 134 – 135) muss eingestehen,
„dass eine detailierte Beschreibung des jeweiligen Merkmals (Lage/Struktur-Kriterien 1 bis 5) und die Feststellung der Merkmalsverbreitung (Merkmalskriterien 6 bis 8) zur Homologieanalyse nicht immer ausreichen. Häufig sind funktionelle und ökologische Kriterien (9 bis 11) für eine detaillierte Analyse essentiell. Leider gibt es erst relativ wenige Fälle, in denen diese Notwendigkeit explizit festgestellt beziehungsweise die verwendeten Ähnlichkeiten auch funktionell/ökologisch detailliert analysiert wurden.“
Nun ist für REMANE (1952) das Problem der Leserichtung dadurch gelöst, dass die Verteilung der homologen Merkmale im System darüber entscheide, in welcher Richtung die Reihen zu lesen sind. So können dann z. B. die Beckenknochen der Wale als Reduktionsreihen gelesen werden, die Zahnformen der verschiedenen Säugetiergruppen hingegen als Komplikationsreihen. PETERS & GUTMANN (1971: 245) kommentieren hierzu:
„Nach den eingangs gemachten grundsätzlichen Überlegungen jedoch ist die Annahme unzutreffend, dass aus der Coracoid-, Becken- oder Zahngestaltung der Säugetiere die Ermittlung der Lesrichtung allein durch Formvergleich möglich sei. In Wirklichkeit sieht REMANE die ‚Säugetiere‘ und ‚Wale‘ seines Beispiels im Gesamtzusammenhang der Wirbeltiere, die schon in phylogenetisch interpretierte Reihen geordnet sind, in denen die Lesrichtung als geklärt angesehen wird. An diese vorgeklärten und zu Stammbäumen zusammengestellten Reihen lassen sich die Beispiele REMANES tatsächlich nur in einer bestimmten Sequenz einfügen. Im Gesamtkomplex der Tetrapoden kann das Becken der Wale nur reduziert sein; die umgekehrte Lesrichtung mit einem progressiven Entwicklungsablauf vom Becken der Wale zu den übrigen Säugetieren passt nicht ins Großdendrogramm der Vertebraten. Unter methodischen Gesichtspunkten bedeutet dies aber, dass die Lesrichtungsentscheidung in die umfassende Problematik der Stellung der Wirbeltiere verlagert ist, dort aber das Problem gar nicht gesehen wird, da offensichtlich die phylogenetische Sequenz der Vertebratenklassen so aussagekräftig angesehen wird, dass eine methodische Erörterung überflüssig erscheint.“
Mit anderen Worten: erst durch die Kenntnis des Stammbaums, also der Evolutionsgeschichte der untersuchten Gruppen, lassen sich Merkmalsreihen lesen. Dies ist im Bereich der Wirbeltiere, vor allem durch ihre relativ gute fossile Überlieferung, die eine zeitliche Reihung der Formen erlauben, Großteils geklärt.
Bei anderen Gruppen sieht die Sachlage jedoch anders aus. Dies zeigte vor allem die „New Animal Phylogeny (kurz: NAP)“ (ADOUTTE et al. 2000), einer mehrfach bestätigten Großphylogenie des Tierreichs basierend auf molekularbiologischen Daten. Das Interessante an dieser molekularen Systematik ist, dass sie mit wesentlichen Lehrmeinungen der traditionellen, morphologischen Systematik in Widerspruch steht. Der bekannteste Widerspruch zeigt die Verwandtschaft der Arthropoden und Anneliden. In der traditionellen Systematik wurden beide Taxa unter dem Großtaxon „Articulata“ zusammengefasst. Als Homologie galt dort u. a. die Segmentierung der beiden Taxa. Die molekulare Systematik hingegen konnte diese Verwandtschaft nicht bestätigen. Die Arthropoden werden zusammen mit den unsegmentierten Nematoden zu den „Ecdysozoa“ zusammengefasst, die neben genetischer Ähnlichkeiten auch das Hormon Ecdyson gemeinsam haben, welches die Häutung bei Arthropoden und Nematoden kontrolliert. Die Anneliden bilden zusammen mit den Mollusken die „Lophotrochozoa“. Gemeinsam ist ihnen neben genetischen Merkmalen, auch die Trochophora-Larve, ein Larvenstadium in der Entwicklung dieser Tiergruppen (vgl. zur Frage der Ecdysozoa vs. Lophotrochozoa-Debatte siehe u.a.: ADOUTTE et al. 2000, AGUINALDO et al. 1997, BALAVOINE & ADOUTTE 2003, BURDA et al. 2008, KUBI 2012, SCHMIDT-RHAESA et al. 1998, SCHMIDT-RHAESA 2004, SYED 2003 & 2006, SYED et al. 2007, WÄGELE et al. 1999).
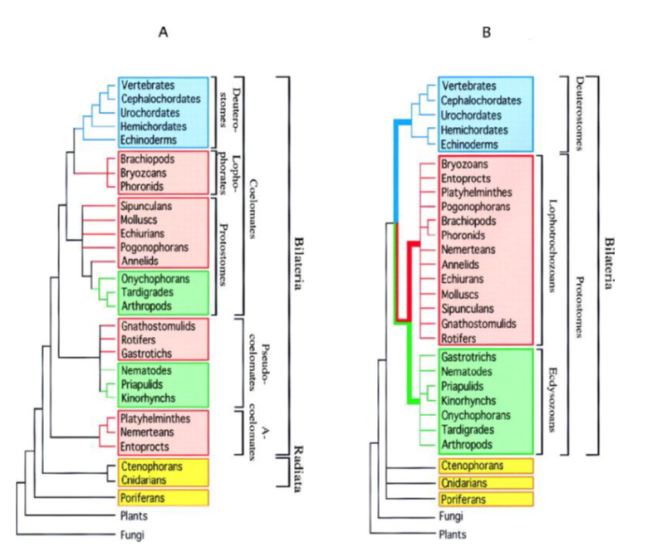
Abb. 8: Phylogenie der Metazoa. (A) traditionelle Systematik gestützt auf morphologische und embryologische Untersuchungen, orientiert am Werk von HYMAN (1940). (B) molekulare Systematik. Bei der molekularen Systematik wird die Monophylie der Deuterostomia (blau) unterstützt, es kommt jedoch zu einer Neuaufteilung der Protostomia in Lophotrochozoa (rot) und Ecdysozoa (grün). Man beachte, dass die Gastrotricha mittlerweile nicht mehr zu den Ecdysozoa sondern zu den Lophotrochozoa gehören. Quelle: ADOUTTE et al. (2000).
Die NAP ist besonders von morphologisch arbeitenden Systematikern nicht unwidersprochen aufgenommen worden. Großteils wurde darauf verwiesen, dass die Methoden der molekularen Systematik unausgereift seien. Tatsächlich konnte WÄGELE (1994) zeigen, dass viele molekulare Analysen unausgereift sind. Dadurch wurde das Vertrauen in die NAP untergraben und die Resultate der NAP wurden von sehr vielen Systematikern abgelehnt (z. B. WÄGELE 1999 et al., WÄGELE & MISOF 2001, SCHOLTZ 2002, SCHOLTZ 2003). Jedoch konnten auch andere molekulare Analysen anderer DNA-Abschnitte die NAP bestätigen (de ROSA et al. 1999, BALAVOINE et al. 2002, GAREY 2003). Desweiteren wurden Fehlerquellen bei der Auswahl genetischer Marker reduziert (ADOUTTE et al. 2000). Besonders erwähnenswert ist jedoch, dass die Ergebnisse der NAP fast alle phylogenetischen Rekonstruktionen der Konstruktionsmorphologie bestätigen, die den merkmalsorientierten Morphologien widersprachen (SYED 2003, 2006). Daher stießen die Ergebnisse der Konstruktionsmorphologie ebenfalls auf Ablehnung.
„Zu betonen ist dabei, dass New Animal Phylogeny und Hydroskelettmodell vollkommen unabhängig voneinander entstanden sind. Für die Rekonstruktion historischer Prozesse – also nicht mehr direkt beweisbarer Vorgänge – gelten aber gerade solche Hypothesen als wertvoll, die von mehreren unabhängigen Datensätzen unterstützt werden.“ (SYED 2003: 67)
Auch wenn die molekularbiologischen Befunde die FAT bestätigen, so kann auch die Molekularsystematik die Konstruktionsmorphologie nicht ersetzen. Es werden lediglich andere Merkmale eingesetzt, die vielleicht auf Grund ihrer Stabilität bei der Replikation geeigneter sind Verwandtschaften aufzuzeigen als die plastischeren morphologischen Merkmale, die mehr Konvergenzen erlauben. Doch auch hier zeigt sich, dass noch längst nicht alle Verwandtschaftsfragen geklärt sind. So gibt es zu den Verwandtschaftsbeziehungen der Euarthropoda mehrere Hypothesen (vgl. BURDA et al. 2008). Desweiteren sollte weiter oben deutlich gemacht worden sein, dass durch die Molekularbiologie Organismen nicht erklärt werden können.
Zwar können mittels molekularbiologischer Vergleiche Verwandtschaften zwischen den Organismen ermittelt werden. Aber ohne konstruktionsmorphologische Erkenntnisse bleiben die molekularen Stammbäume leere Schemata (GUTMANN 1995b). Aussagen zur Formveränderung, zur Anagenese, können in der phylogenetischen Systematik nicht gemacht werden. Um hier einer Analogie gerecht zu werden: mit einem Vaterschaftstest sind sehr sichere Aussagen zu machen, wer der Vater eines Kindes ist oder nicht. Über den Prozess der Zeugung, der Embryonalentwicklung und der Geburt werden jedoch keine Aussagen getroffen.
„Wie sinnvoll eine kladistische Aussage ist, hängt von der Art der Argumentation ab. Verweist man auf eine Baueigenheit, die man rekonstruieren konnte, so ist das wissenschaftstheoretisch in Ordnung, (…). Legt man aber nur idealistisch-morphologische gewonnene Merkmale zu Grunde, so überschreitet man die Grenze des Geltungsbereiches der beschreibenden Morphologie. Eben letzteres wird heute in der Kladistik praktiziert: Indem man viele Merkmale einbezieht, erstellt man umfangreiche Diagramme sich dichotom verzweigender Linien für eine Tier- oder Pflanzengruppe. Es wird heute kaum noch eine taxonomisch beschreibende Arbeit ohne solche ‚phylogenetische Analyse‘ zur Publikation in einer Fachzeitschrift angenommen. Vor allem in Kombination mit Molekularbiologie gilt Kladistik heute als einzig legitime Methode des Fachgebiets. Die so erstellten Stammbäume zeigen nichts anderes als Merkmals-Vergleiche. (…) Man versucht homologe Organe, Homologien genannt, durch idealistisch-morphologischen Vergleich zu ermitteln, woraus man dann auf Stammesverwandtschaft schließt.“ (Grasshoff 2014)
Offen bleibt jedoch noch die Frage wie Lesrichtungskriterien ermittelt werden sollen. In der Kladistik, die methodisch wie oben gezeigt wurde keine Kriterien hierfür besitzt, setzt sich zunehmend ein Parsimonieprinzip durch. Da apomorphe und plesiomorphe Merkmale nach dem Wahrscheinlichkeitsprinzip festgelegt werden, geht man auch von einem möglichst sparsamen Bauplan aus. Dies zeigt sich z. B. in der Frage, ob der Urbilateria segmentiert war oder nicht. BALAVOINE & ADOUTTE (2003) gehen aufgrund der Befunde der NAP davon aus, dass der Urbilateria auch segmentiert sein musste. Die traditionelle morphologisch Systematik ging aufgrund der postulierten Verwandtschaft der Arthropoden und Anneliden davon aus, dass der Urbilateria unsegmentiert war. So argumentiert u. a. SCHMIDT-RHAESA (2004: 44 f.):
„Wenn die Segmentierung bei der Stammart der Bilateria evoluierte, dann muss sie in vielen getrennten Linien verloren gegangen sein. Rekonstruktionen des Grundmusters der Bilateria aufgrund des Sparsamkeitsprinzips lassen daher eine segmentierte Stammart als unwahrscheinlich erscheinen.“
SYED (2006: 34) entgegnet hierzu:
„Das eigentliche Hauptargument aber ist … eine reine Parsimoniebegründung. Demzufolge müßten numerisch betrachtet mehr Bilateria-Phyla die Segmentierung sekundär verlieren, als sie zu behalten. Deshalb wiederum sei es wahrscheinlicher, eine mehrfach unabhängige Entstehung der Segmentierung und eine konvergente Evolution der genetischen Segmentationsmechanismen anzunehmen.“
Doch auch diese numerische Parsimoniebegründung führt ins Leere, da auf morphologischer Ebene keine Begründung für den Verlust der Segmentierung gesucht wird (SYED, GUDO & GUTMANN 2007). GUTMANN (1972, 1995) konnte schon konstruktionsmorphologisch aufzeigen, dass der Urbilateria segmentiert gewesen sein musste. Konstruktionsmorphologisch betrachtet ist es das schon oben beschriebene Ökonomieprinzip der phylogenetischen Systematik, dass die Ermittlung phylogenetischer Reihen durch die Rekonstruktion des Ausgangsprozesses bestimmt (PETERS & GUTMANN 1971). Dabei ist unter den Ökonomieprinzip nicht ein Abzählen plesiomorpher oder apomorpher Merkmale gemeint. Durch die Konstruktionsbedingungen sind in der Entwicklung der Organismen nur begrenzte Bahnen der Entwicklung möglich, die in klar begründbarer Abfolge ermittelt werden müssen (GRASSHOFF 1994a).
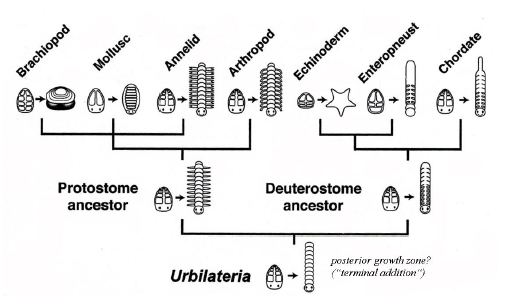
Abb. 9: Stammbaum der Bilateria leicht verändert nach BALAVOINE & ADOUTTE (2003). Der Urbilaterier wird als segmentiert dargestellt.
Organismus Anpassung und Selektion
Die Grundlagen der Konstruktionsmorphologie führen auch zu einer Revision des Evolutionsdenken. Evolution wird hauptsächlich so verstanden, dass diese als ein Prozess der Anpassung der Organismen an ihre Umwelt gesehen wird (VOGEL 1991). Organismen werden also gar nicht aus sich selbst heraus, sondern als Abbild der Umwelt verstanden (GUTMANN & BONIK 1981). Der Ansatz der Konstruktionsmorphologie stellt jedoch die Organismen in das Zentrum des Evolutionsdenkens (BONIK et al. 1984). Es wird also die aktive Rolle der Organismen stärker betont und als Subjekt in Bezug auf seine Umwelt gesehen. Was für einen Organismus Umwelt ist, ergibt sich aus seiner internen Struktur und Organisation. Seine Organisation bestimmt auch in welche Umwelt ein Organismus vordringen kann (EDLINGER et al. 1991). Das heißt, dass die Aspekte der Internselektion in den Vordergrund gerückt werden, da Wandel in der Umwelt nur nach Maßgabe der internen Strukturierung und des ungehinderten Energiewandels geschehen kann. Dabei werden in den Rekonstruktionen der Konstruktionsmorphologie anders als z. B. REMANE (1983) behauptet, der Umweltbezug nicht ignoriert:
„Der Umweltbezug jedes Lebewesens ist natürlich immer nötig. Das System muss Material und Energie aus der Umwelt aufnehmen. Es muss seine Leistung in der Umwelt erbringen. Dass es dies kann, ist aber nur vom Verständnis des Systems aus möglich und nicht von der Umwelt her zu erschließen. Außerdem können Systeme nach Ausbildung bestimmter Einrichtungen, die vorher in anderen Bereichen Vorteile brachten, in neue Lebensräume vordringen. Das heißt Organismen überleben nach Maßgabe ihres Aufbaus auch in anderen Bereichen. Dies führt aber jede Vorstellung einer Erklärung der Organismen von der Umwelt her ad absurdum. Will man begründen, warum ein Organismus in einem Lebensrum lebt, so muss man die Erklärung aus der Leistung des Apparates ableiten; auch kann man nur vom Apparat her verständlich machen, in welche neuen Lebensräume er eindringen kann.“ (GUTMANN 1980: 12)
Organismen sind also durch die Stoffwechselerfordernisse von der Außenwelt abhängig und damit nicht total autonom. Aber was Organismen als Umweltbedingungen nutzen können, lässt sich nur an der organismischen Konstruktion bestimmten (EDLINGER et al. 1991).
Ob also ein Organismus in seiner Umwelt überleben und sich reproduzieren kann, hängt also von seiner Konstruktion ab (GUTMANN 1989, 1995, GUDO 1999, 2007). Es werden nämlich diejenigen Nachkommen selektiert, die nicht in der vorliegenden Umwelt zurechtkommen. Teilphasen der rekonstruierten Abläufe laufen ohne Umweltwechsel, aber immer mit Umweltbezug (BONIK et al. 1981). Es findet also Evolution auch ohne Umweltänderung statt, also auch wenn Ressourcen im Überfluss vorhanden sind (GUTMANN 1980). Grund hierfür ist, dass die Ursachen der Evolution im Energiewandel der lebenden Konstruktionen zu suchen ist und nicht in den Veränderungen der Umweltbedingungen. Entsprechend ist die vom Organismus festgelegte Umwelt nicht mit der darwinistischen Milieu-Vorstellungen zu vergleichen, sondern es ist der Organismus der seine Umweltbedingungen erschafft und die Umwelt als Energie- und Materialquelle nutzt, sich ihrer Widerständigkeit bei der eigenen Aktivität bedient und seine Freiräume schafft (EDLINGER et al. 1991). Dabei wird jedoch nicht ignoriert, dass es Teilorgane oder Strukturen gibt, die sich als direkte Anpassungen an die Umwelt darstellen (BONIK et al. 1984). Es wird auch nicht geleugnet, dass das Außengeschehen der Umwelt (ob abiotische oder biotische Faktoren) auf die Organismen einwirken, ihn variieren und zerstören können. Wirken sie nicht zerstörend, erfolgt ihre Wirkung auf die Konstruktion (EDLINGER et al. 1991). Der Organismus in seinem Kern aber ist von einer Anpassung durch die Umwelt nicht erkennbar, da er einen Stoffwechsel, eine Biomechanik und damit eine erkennbare, wenn auch nicht totale, Autonomie hat (BONIK et al. 1984). Anpassung ist dabei als Eingliederungsfähigkeit der Organismen zu verstehen. Wenn neue Varianten sich in der Umwelt zurechtfinden, also an diese adaptiert sind, wird diese schon festgelegt bevor, die Selektion eingreifen kann (GRASSHOFF 1994). In populärwissenschaftlichen Darstellungen oder in Lehrbüchern wird oft das Bild des besser oder schlechter Angepassten vermittelt. Tatsache ist jedoch, dass in der gleichen Umgebung und Umweltbedingung verschiedene Konstruktionen vorkommen und bei gleichen Umweltbedingungen können verschiedenartige Entwicklungen von gleichen Vorentwicklungen möglich sein (EDLINGER et al. 1991). Dabei ist die Anpassung stets relativ zu sehen. Es gibt keine Anpassung schlecht hin, sondern immer in Bezug auf eine Konstruktion, zumal kein Organismus auf nur eine Funktion ausgerichtet ist (PETERS et al. 1971). Beispielsweise sind wir Menschen zwar überwiegend bipedale Bodenläufer. Das hindert uns jedoch nicht daran zu klettern, schwimmen oder zu robben. Das Problem der Definition der Anpassung ist, dass dieser so weit gefasst ist, dass er auf alle Phänomene angewandt und ausgedehnt werden kann. Somit wird alles und nichts zugleich erklärt. Selbiges gilt für Selektion. Beide Begriffe sind als ex-post-Aussagen immer richtig, da festgestellt wird, dass die Überlebenden überleben, weil sie angepasst sind (GUDO 2007). Organismen die nicht an ihren Lebensraum angepasst sind gibt es von vornherein nicht, da sie schon vor ihrer Entstehung ausselektiert werden.
Der Anpassungsbegriff im Darwinismus ist analog zur Züchtungspraxis entworfen worden. Die natürliche Auswahl durch die Umwelt entsprach so dem Bilde eines auswählenden Züchters. Die Wirkung der Selektion liege so außerhalb des Organismus, der nur Objekt im Anpassungsprozess sein konnte. Dabei ist das Sieb eine Metapher der Selektion, mit dessen Hilfe man Partikel nach ihrer Größe sortieren kann. Jedoch liegt die Ursache der Partikelgröße nicht am Sieb, sondern an den Partikeln selbst. So kann man organismische Konstruktionen ebenfalls nicht durch den Selektionsprozess, sondern wenn man sich mit ihnen selbst befasst. Organismen leben zwar in einer Umwelt und sind durch den Stoffwechsel auf diese angewiesen. Aber die Organismen bestimmen durch ihre Konstruktion, was die ihnen angemessene Umwelt ist (PETERS & PETERS 1995). Dabei ist diese Tatsache nicht teleologisch zu verstehen, sondern ist die notwendige Folge ihrer Konstruktion. Organismen nehmen Energie aus der Umwelt auf, wandeln diese um und betreiben die Konstruktion. Schon GOULD & LEWONTIN (1979) machten in ihrem Essay „The Spandrels of San Marco and the Panglossian Paradigm: A Critique of the Adaptationist Programme“ deutlich, dass nicht alle Merkmale adaptiv sind, sondern konstruktionsbedingt vorhanden sein müssen. Besonders in entwicklungsbiologisch orientierten Publikationen ist zunehmend von „constraints“ die Rede (z. B. ALBRECH 1989, FORGACS & NEWMAN 2005, GILBERT & EPEL 2009), die klar darstellen, dass es wichtige konstruktionsbedingte Grenzen in der Entwicklung gibt, auch wenn allen Autoren gemein ist, dass sie Organsimen nicht als energiewandelnde hydraulische Einheiten beschreiben.
Zum anderen werden zunehmend Organismen auch nicht als rein passive Wesen, sondern als aktive Gestalter gesehen, die ihre Umwelt und die anderer Organismen erschaffen (sog. Niche construction, siehe LALAND et al. 2008)
GRASSHOFF (2014) macht deutlich, dass der Begriff Spezialisierung aussagekräftiger ist als der Begriff Anpassung. So sei ein Thunfisch an das offene Wasser angepasst, wie viele andere Konstruktionen auch, die jedoch ganz anders aussehen (z. b. Robben, Plesiosaurier, Meeresschildkröten). Wenn man jedoch sagt, ein „Thunfisch ist mit seinem Bewegungsapparat auf schnelles Schwimmen und Jagen im offenen Ozean spezialisiert und der Rotfeuerfisch auf kurzes Rudern und Wendigkeit zwischen den Korallenblöcken und ruckartiges Zuschnappen nach Beute“, so erhellt das die Beziehung von Körperkonstruktion und Umwelt. Denn die Rekonstruktion der selektierenden Einflüsse der Umwelt kann nur gelingen, wenn die organismische Struktur und die Umwelt in Rechnung gestellt werden (PETERS & PETERS 1995). Dabei muss der Begriff der Selektion um den Aspekt der internen organismischen Bedingungen erweitert werden (GRASSHOFF 2014).
Die Konsequenzen dieser Erkenntnis reichen jedoch noch weiter:
Als Energiewandler bestimmt die Körperkonstruktion welche Energie von außen aufgenommen, im Inneren umgewandelt und nach außen abgegeben wird. Organismen wirken aktiv auf die Umwelt ein, indem er Energie wandelt und mechanische Arbeit leistet. Nach Maßgabe ihrer Konstruktion dringen Organismen in die Umwelt vor und verändern sie mit (EDLINGER et al.).
„Die Folge der Ausbreitung ist, dass sich diese Populationen unterschiedlich auf Nahrungserwerb und Raumnutzung spezialisieren. Die damit Erschlossenen Lebensräume werden zur ‚Umwelt‘ der jeweiligen Organismen. Daraus wird deutlich, dass Organismen in Kohärenz mit ihrer spezifischen Umwelt stehen, ganz entsprechend der Kohärenz der Teile in ihrem Inneren. Die Spezialisierung sorgt sozusagen für einen günstigen externen Wirkungsgrad.“ (GRASSHOFF 2014: 69)
Organismen dringen nicht nur nach Maßgabe ihrer Konstruktion in neue Lebensräume ein, sondern gestalten sie auch maßgeblich mit. Organismen sind in den Energiefluss, die von der Sonne ausgeht eingeklinkt, indem von einigen (den Produzenten) Photosynthese betrieben wird. Die Sonnenenergie wird in andere Energie umgewandelt und für andere Organismen nutzbar gemacht. Es wird somit die Sonnenenergie in die Biosphäre eingespeist. Somit sind Biosphäre, Atmosphäre und Hydrosphäre in ein organismisch gesteuertes Gesamtsystem integriert und Organismen erzeugen so ihre eigenen Lebensräume (GUTMANN & EDLINGER 1994, GUTMANN 1996a, b, 1997, GUDO 2007). Die Folge dieser Prozesse ist, dass Organismen ständig neue thermodynamische Entropiezustände erzeugen und aufnehmen und sich über ich generationenübergreifendes Daseins ständig wandeln. Evolution ist damit notwendigerweise das Resultat der Organismen als Energiewandler (GUDO 2007).
„Lebewesen existieren in den Materie- und Energieströmen der Biosphäre, die wie eine durch die Energie der Sonne betriebene Megamaschine arbeitet. Organismen sind aber nicht nur in die Materieströme eingeflochten, denn sie erzeugen (…) Eigenheiten der Außenwelt selbst und regulieren sie im hohen Maße. Die Atmosphäre, Hydrosphäre und die Ausgestaltungen der Erdablagerungen, der Riffe und der Stromatolithen wird durch Organismen erzeugt und/oder hochgradig reguliert“ (GUTMANN 1997: 125)
Diese Vorstellungen einer von Lebewesen gestalteten Erde wird auch in den Arbeiten LOVELOCK (1988) und MARGULIS (1998), MARGULIS & DOLAN (2002) und anderen Arbeiten dieser Autoren als GAIA-Hypothese ausgearbeitet.
Nach GUTMAN & EDLINGER (1994: 15) sind auch Lebens- und Erdentwicklung voneinander unabhängig:
„Auf organismischer Grundlage stellt sich Evolution als ein geschehen dar, das die Erde als Unterlage nutzt, jedoch als eigenständiges Geschehen durchläuft. Evolution von Organismen geschieht autonom, also nicht nach dem LYELLSCHEN Prinzip, nach dem die geologische Entwicklung die Evolution der Organismen treiben und richten soll. Geologisches Geschehen läuft im Sinne der Plattentektonik als eigengesetzliches Geschehen ab, ist ebenfalls autonom, vermag auch Evolution nicht zur richten oder in der Abfolge der Stadien zu bestimmen. Bei aller Dynamik des geologischen Geschehens blieben doch die generellen Bedingungen des Geschehens seit mehr als einer Milliarden Jahre invariant, zeigten also im Wandel jederzeit gleiche Bedingungen. Indem in jedem Zeitschnitt alle Grundbedingungen gegeben sind, kann das Leben sich im Wandel die ihm passenden Situationen für die intern bedingte Weiterentwicklung suchen. Durch die geologische Situation werden eventuell Entwicklungen abgeschnitten, die dann andernorts weiterlaufen, oder an manchen Stellen Entwicklungen versperrt, während sich in anderen geographischen Bereichen die Weiterentwicklung vollzieht. Es kann geologisch nicht die Entwicklung der Organisation oder das Unterbleiben anderer Entwicklungen erklärt werden, auch die Negativ-Form des Lyell-Prinzips gilt nicht.“
Das Evolutionsgeschehen verläuft also nach eigenständigen Prinzipien. Durch diese Einsicht erlangt die Geologie aber auch keinerlei Schaden, sie wird tatsächlich in einem Punkt dynamisch weiterentwickelt. Denn durch Organismen werden auch geologische Prozesse erzeugt. Z. B. sind Riffe Ergebnis organismischer Tätigkeit. Organismen wirken gestaltend und teils auch zerstörend auf die Gestalt der Erdoberfläche ein. Durch die Kontinentalbewegung (Plattentektonik), die auch unabhängig der Tätigkeit der Organismen läuft, werden diese Bildungen mitgenommen. So können z. B. aus ehemaligen Riffen (also dem Produkt organismischer Leistung) durch Verschiebung der kontinentalen Platten Gebirge entstehen (GUTMANN & EDLINGER 1994).
Dieser Theorie nach haben Fossilien für die Paläontologie auch eine andere Bedeutung. Durch Fossilienreihen kann wie durch die oben beschriebenen Homologiereihen kein Evolutionsverlauf erklärt werden:
„Dies bedeutet nun nicht, dass fossile Funde nicht wichtige Dokumente sein könnten. Das Gegenteil ist der Fall. Nur Fossilfunde gestatten die zeitliche Einordnung, nur Funde belegen, wann eine rekonstruierte Entwicklung im geologischen Rahmen abgelaufen ist. Radiationen auf verschiedenen Niveaus lassen sich fossil untermauern und dokumentieren, begründbar sind sie nur über die vorgeordneten konstruktiven Einsichten. Bei skeletttragenden Formen können auch verschiedene Entwicklungsstadien, die aus der Modellbegründung gefordert werden, Stufen des Evolutionsprozesses belegen; Voraussetzung für solche Erklärungen ist aber immer, dass die Skelette oder Skeletteile als Elemente lebender Konstruktionen erklärbar und so auch nutzbar sind.“ (GUTMANN & EDLINGER 1994: 18).
Abschließende Betrachtungen
Die Konstruktionsmorphologie nach Gutmann liefert wertvolle Erkenntnisse über Bauplan und Evolution der Organismen, insbesondere der Tiere. Sie hat es geschafft über die Evolution der Tiere eine gewisse Klarheit zu schaffen, ohne den Organismus dabei aus den Augen zu verlieren. Die in der klassischen Evolutionsbiologie hilfreichen Vergleiche der Merkmale und Homologien haben zum Nachteil, dass man bei der Fülle der Merkmale und der Vielfalt der Organismen das Tier vor lauter Merkmalen nicht mehr sieht. Dies führte dazu, dass – besonders bei den vielfältigen oder uns Menschen als „Gewürm“ bezeichneten Tiergruppen wie Insekten, Weichtiere, Korallen und Schwämme –verschiedene Stammbäume und Verwandtschaftsgrade der Organismen erstellt wurden, ohne dabei die Funktionalität, Einheit und Lebensfähigkeit des Gesamt-Organismus zu berücksichtigen. Die Konstruktionsmorphologie schaffte es die wesentlichen Eigenschaften aller (tierischen) Organismen – Hydraulik, Energiewandel, Kohärenz etc. – herauszuarbeiten und basierend auf diesen Prinzipien eine plausible Evolutionsgeschichte der Organismen zu erstellen. Diese stand teilweise im krassen Gegensatz zu den rein auf morphologischen Merkmalen basierenden Stammbäumen. So erkannte die Konstruktionsmorphologie, dass der Ur-Bilaterier schon segmentiert gewesen sein muss und dass die unsegmentierten Nematoden den segemtierten Arthropoden näher stehen, als zu den ebenfalls segmentierten Annelida. All diese Stammbäume der Konstruktionsmorphologie basierten auf die Funktionsweise der Organismen, die nicht einfach so einzelne Merkmale hinzuaddieren können, da es aus konstruktionsmorphologischer Sicht energiezehrend – und damit evolutionär nachteilig – wäre. Dabei sei angemerkt, dass es sich schwerpunktmäßig um die Großgruppen im Tierreich (also um die Tierstämme und Klassen) handelt und nicht um Aufspaltungen in unterschiedliche Arten. Bemerkenswert ist, dass die Einführung der auf molekularen Merkmalen basierenden Stammbäume zu ebenso widersprüchlichen Aussagen mit den auf morphologischen Merkmalen basierenden Stammbäumen führte. Auf der anderen Seite fanden sich viele Übereinstimmungen mit den konstruktionsmorphologischen Überlegungen. Die Konstruktionsmorphologie schaffte es also die Kluft zwischen den verschiedenen Methodenansätzen der Phylogenie zu vereinen und bestehende Widersprüche und Streitigkeiten zu lösen.
Eine weitere Stärke der Konstruktionsmorphologie ist auch, dass sie den Organismus vor reduktionistischem Denken, welches den Organismus nur als Träger von Genen sieht, zu befreien.
Durch ihre gänzlich andere Herangehensweise an die Morphologie schossen die Theorien Gutmanns jedoch auch weit übers Ziel hinaus. Statt sich ergänzend in den Wissenschaftsstreit in der Morphologie einzubinden, wollte man ‚altdarwinistische Dogmen‘ über Bord werfen und nur die konstruktionsmorphologischen Prinzipien als Evolution anerkennen. Über Selektion und Anpassung, wie auch der Artenbildung zu spekulieren, sei nach Gutmann keine richtige Evolutionsforschung. Das stieß verständlicherweise unter den Biologen auf Unverständnis mit dem erheblichen Nachteil, dass die Konstruktionsmorphologie zu einer kaum anerkannten – und über dem deutschsprachigen Raum praktisch unbekannten – wissenschaftlichen Theorie und Methode wurde. Gleich ideologischen Debatten politischer Sekten des maoistischen oder trotzkistischen ultra-linken Spektrums (Volksfront von Judäa vs. Judäische Volksfront) wurde beiderseits mit Strohmann-Argumenten, persönlichen Diffamierungen und Ausgrenzungen gearbeitet, was der Konstruktionsmorphologie mehr Schaden zufügte, als Nutzen.
Auch wenn das Anpassungsdenken der Konstruktionsmorphologie dem der klassischen – darwinschen – Evolutionstheorie als Gegenpol erscheint, so stehen sie nicht unbedingt im antagonistischen Widerspruch zueinander. Es handelt sich halt eben um zwei verschiedene Betrachtungsweisen desselben Phänomens: Während die Konstruktionsmorphologie den Organismus in den Mittelpunkt stellt, stellt die klassische Evolutionstheorie die Wirkungen der Umwelt in den Vordergrund. Da die Konstruktionsmorphologie weder die Abhängigkeit des Organismus von seiner Umwelt leugnet, noch die klassische Evolutionstheorie den Organismus als passives Opfer der Umweltbedingungen sieht, handelt es sich lediglich um zwei verschiedene Konzepte der Beschreibung der Evolution. Organismen stehen in einem dialektischen Wechselwirkung mit ihrer Umwelt und ihrer Konstruktion. Ein Organismus wandert nach Maßgabe seiner Konstruktion in die Lebensräume ein und nutzt die nach seiner Konstruktion entsprechenden Energiequellen. Die Umwelt aber liefert eben diese Energiequellen und wenn diese fehlen, hilft die beste Konstruktion nichts. Außerdem basiert ja die Konstruktion der Organismen selbst auf Naturgesetze (Hydraulik, Mechanik, Thermodynamik), womit sie Teil der Umwelt selbst sind.
Und je nachdem welcher Fragestellung man hat, eignet sich die eine oder die andere Theorie zu dessen Beantwortung. Entsprechend wenden beide unterschiedliche Methoden zur Beantwortung ihrer Fragen an und beide haben eine Daseinsberechtigung. Eigentlich sollten sich beide Konzepte eher einander Ergänzen als Konkurrieren. Das kann anhand eines Beispiels erklärt werden, nämlich der Mimikry bei Insekten.
Unter Mimikry ist die (farbliche oder formliche) Nachahmung einer Art durch eine andere zu verstehen. Beispielsweise ahmen ungiftige, harmlose Tiere durch ihre Färbungen giftige Tiere nach. So ist von vielen harmlosen Schmetterlingsarten bekannt, dass sie durch ihre Farbmuster auf den Flügeln giftige, ungenießbare Schmetterlinge meiden. So sind sie vor Fressfeinden geschützt und können sich erfolgreich reproduzieren. Aus Sicht der klassischen Evolutionstheorie haben wir hier einen klaren fall der Evolution: Arten verändern sich (harmlose Arten verändern ihr Farbmuster), passen sich ihrer Umgebung (giftige Arten) an sichern so ihr Überleben und zeugen mehr Nachkommen (durch Vermeidung von Fressfeinden). Aus Sicht der Konstruktionsmorphologie handelt es sich jedoch bei beiden Arten um die gleiche Arthropoden-Konstruktion, es bestehe also kein wesentlicher Unterschied zwischen der giftigen und ungiftigen Art, sondern lediglich Variationen der selben Konstruktion. Während der Arbeitsgegenstand der klassischen Evolutionstheorie Arten, Populationen, Fortpflanzungsgemeinschaften oder gar Individuen sind, beziehen sich die Aussagen der Frankfurter Evolutionstheorie auf die den jeweiligen Lebewesen eigenen Körperkonstruktionen. Eine Konstruktion ist nicht gleichzusetzen mit einer taxonomischen Kategorie. Organismische Konstruktionen sind vielmehr Beschreibungsweisen von Lebewesen hinsichtlich ihres Aufbaus und Funktionierens.
Beides beschreibt Evolution, hat aber verschiedene Anknüpfungspunkte.
Nun gut 30 – 40 Jahre nach diesen heftigen Debatten, die keineswegs eine Seltenheit in wissenschaftlichen Kreisen sind, wenn neue Ideen hervorgebracht werden, sollte es wieder an der Zeit sein, sich die positiven Seiten der Konstruktionsmorphologie zu eigen zu machen. Es wird und wurde oftmals außer Acht gelassen, dass sowohl Konstruktionsmorphologie als auch der klassische Darwinismus nicht in Widerspruch zueinander stehen müssen, sondern sich wunderbar ergänzen können, wie es sich anhand der molekularen Stammbaums der Tierstämme zeigt. Beide Theorien haben in unterschiedlichen Bereichen einen Geltungsanspruch: Während die Konstruktionsmorphologie sich gut dafür eignet makro-evolutionäre Prozesse begreiflich zu machen, hat der klassische Darwinismus seine Stärken in mikro-evolutionären Prozessen (natürliche und sexuelle Selektion, Artenbildung, Veränderungen in Populationen etc.). Wir haben hier eine dialektische Einheit der Gegensätze, wo sich zwei konkurrierende Denkmodelle dadurch vereinen, dass sie unterschiedliche Ansatzpunkte in der Evolutionsforschung haben und diese sich wunderbar ergänzen können, da sie damit auch unterschiedliche Qualitäten des biologischen Lebens erforschen. Es bleibt zu hoffen, dass sich diese Widersprüche und Missverständnisse in Zukunft lösen.
