

*Anmerkung: dieser Artikel basiert z. T. auf den Artikel „Lies, Damned Lies, Statistics, and Probability of Abiogenesis Calculations“ von Ian Musgrave aus dem Jahr 1998, erschienen auf der Homepage von talk-Origins[1]. Entsprechend haben die hier zitierten Quellen ein entsprechendes Alter. Hinzugefügt wurden weitere Abschnitte, die sich mit der Laborarbeit allgemein und Thermodynamik auseinandersetzen. Die Grundlagen zur Thermodynamik finden sich in jedem Standard-Lehrbuch der Physik oder Chemie, weswegen hier nicht auf entsprechende Quellen verwiesen wird. Aber eine aktuelle Auseinandersetzung mit dem Kreationismus und ihren mathematischen und thermodynamischen Pseudoargumenten findet sich in „The Failures of Mathematical Anti-Evolutionism“ von Jason Rosenhouse (2022).[2] Zwischen diesem Buch und dem für diesen Artikel verwendeten Grundlagentext liegen über 20 Jahre. Doch ein genauer Blick verrät, dass die Argumente der Evolutionsgegner praktisch dieselben sind, lediglich mit anderen Beispielen unterfüttert. Daher büßt der Artikel von Ian Musgrave nichts an Aktualität ein. Weitere biologische Grundlagen und wissenschaftliche Arbeiten zur Abiogenese sind entsprechend in dieser Serie zur Entstehung des Lebens zu entnehmen.
Text als pdf
Es kommt immer wieder vor, dass jemand behauptet: „Die zufällige Bildung eines Proteins, Gens etc. ist nahezu unmöglich, also ist auch die Entstehung des Lebens durch ‚Zufall‘ unmöglich“.
Oft kommen sie mit beeindruckenden Berechnungen zur Unwahrscheinlichkeit oder kommen mit erstaunlichen Aussagen, dass die natürliche Entstehung des Lebens unmöglich ist.
So sagte Fred Hoyle: Die Wahrscheinlichkeit, dass Leben auf der Erde entsteht, ist nicht größer als die, dass ein Wirbelsturm, der über einen Schrottplatz fegt, rein zufällig eine Boeing 747 zusammenbaut (Abb. 1).

Abb. 1: Behauptung von Fred Hoyle
Die Begründung ist wie folgt:
Es gibt 20 Aminosäuren, die in jeder beliebigen Kombination angeordnet werden können, und ein durchschnittliches Protein besteht aus über 300 miteinander verbundenen Aminosäuren. Die Eigenschaften und die Funktion des Proteins werden durch die Anzahl und die besondere Anordnung der Aminosäuren bestimmt. Dazu kommt noch das Problem der Chiralität, so gibt es linkshändige und rechtshändige Aminosuren, Lebewesen nutzen aber nur die linkshändige Form. Es sei mathematisch unmöglich, dass sich zufällig die 300 richtigen Aminosäuren miteinander verbinden, um ein funktionierendes Protein zu bilden.
Die zufällige Bildung von funktionalen Proteinen ist vergleichbar mit einem Affen, der versucht, eine Seite Shakespeare mit den 26 Buchstaben des Alphabets zu schreiben. Jeder weiß, dass der Affe nicht in der Lage ist, die ihm gestellte Aufgabe zu bewältigen.
All diese Beispiele werden als Dilemma der Evolutionstheorie dargestellt und zugleich als Beweis dafür, dass ein intelligenter Schöpfer dahinterstecken muss. Doch das Problem ist, dass bei all diesen „Unwahrscheinlichkeitsannahmen“ gravierende Denkfehler vorhanden sind. Ich werde versuchen, diese verschiedenen Fehler zu erklären und zu zeigen, warum es nicht möglich ist, eine sinnvolle Berechnung der „Wahrscheinlichkeit der Abiogenese“ durchzuführen.
1. technische Objekte sind keine biologischen
So dumm das auch klingt, dieses Argument wird überall verwendet, von wissenschaftlich ungebildeten Predigern bis hin zu anerkannten Befürwortern des Intelligent Design.
Erstens ist der Vergleich des Lebens mit einem vom Menschen geschaffenen Objekt ebenso absichtlich wie absurd. Es ist bekannt, dass von Menschen geschaffene Objekte menschliche Designer haben. Natürlich setzen sie sich nur so zusammen, wie wir sie nach unserem Entwurf zusammengesetzt haben. Das hat absolut nichts mit der Natur zu tun.
Zweitens hat diese Vorstellung, dass man einfach Teile zusammenklebt, um ein Objekt zu schaffen, wiederum sehr wenig mit dem zu tun, was die Wissenschaft vorschlägt. Ein Prozess, der eindeutig sowohl iterativ als auch graduell war, kann nicht mit einem singulären Ereignis verglichen werden. Generell ist jede Analogie, die sich auf vom Menschen geschaffene Objekte bezieht, von Natur aus irreführend. Die Implikation ist, dass Menschen komplexe Dinge erschaffen, also muss alles, was komplex ist, von jemandem erschaffen worden sein. Dies fällt sofort auseinander, wenn man die Natur betrachtet. Schneeflocken sind geometrisch hochkomplex, so dass ihre Schönheit mit der von Menschen geschaffenen Kunst und Architektur konkurrieren kann. Sie entstehen jedoch durch rein natürliche Prozesse in der Atmosphäre.
Letztendlich korrelieren Komplexität und Design überhaupt nicht miteinander. Das wird überdeutlich, wenn wir akzeptieren, dass Komplexität nicht das Kennzeichen von Design ist. Es ist die Einfachheit. Ein iPad hat nicht Tausende von Tasten, von denen einige mehr als eine Funktion haben, einige die gleiche Funktion wie andere und einige gar keine haben. Es hat eine einzige Taste. Er ist so konzipiert, dass er einfach und intuitiv zu bedienen ist. In der biologischen Welt ist das ganz anders. Die Natur plant nichts. Sie probiert alles aus und behält, was funktioniert. Es gibt kein Ziel, es gibt keinen Bauplan, keine Funktion ist vorherbestimmt, man stolpert darüber. Ohne Empfindungsvermögen verlässt sich die Natur ausschließlich auf Mutation und Selektion.
2. Eine protoplasmatische Urkugel
Die Berechnungen der Evolutionsgegner sagen, dass es sehr unwahrscheinlich sei ein funktionierendes Protein aus sagen wir mal 300 Aminosäuren (das entsprecht in etwa der Länge des Enzyms Carboxypeptidase) zu bilden. Die Wahrscheinlichkeit betröge in diesem Fall (1/20)300 oder 1: 2,04 x 10 390 – definitiv eine astronomisch hohe Zahl (Abb. 2). Da eine durchschnittliche Zelle natürlich mehr als nur einen Typ von Proteinen besitzt, werden die Zahlen so groß, dass einem schon bei der bloßen Betrachtung das Gehirn aus den Ohren tropft. Das erweckt den Eindruck, dass die Entstehung auch nur eines kleinsten Organismus völlig unmöglich erscheint. Dies ist jedoch völlig falsch.

Abb. 2: Wie wahrscheinlich ist es, dass sich ein Protein aus 300 Aminosäuren bilden kann?
Erstens unterliegt die Bildung biologischer Polymere aus Monomeren den Gesetzen der Chemie und Biochemie, und die sind keineswegs zufällig.
Zweitens ist die gesamte Prämisse von vornherein falsch, denn in modernen Abiogenese-Theorien wären die ersten „Lebewesen“ viel einfacher, nicht einmal eine Protobakterie oder eine Präprotobakterie (was Oparin als Protobiont und Woese als Progenote bezeichnet; Ruse 1997, Woese 1998), sondern ein oder mehrere einfache Moleküle, die wahrscheinlich nicht länger als 30-40 Untereinheiten sind. Diese einfachen Moleküle entwickelten sich dann langsam zu kooperativeren selbstreplizierenden Systemen und schließlich zu einfachen Organismen (Orgel 1998, Varetto 1998, Lee et al. 1997a, Lazcano & Miller 1996, Eigen & Schuster 1979; Abb. 3).
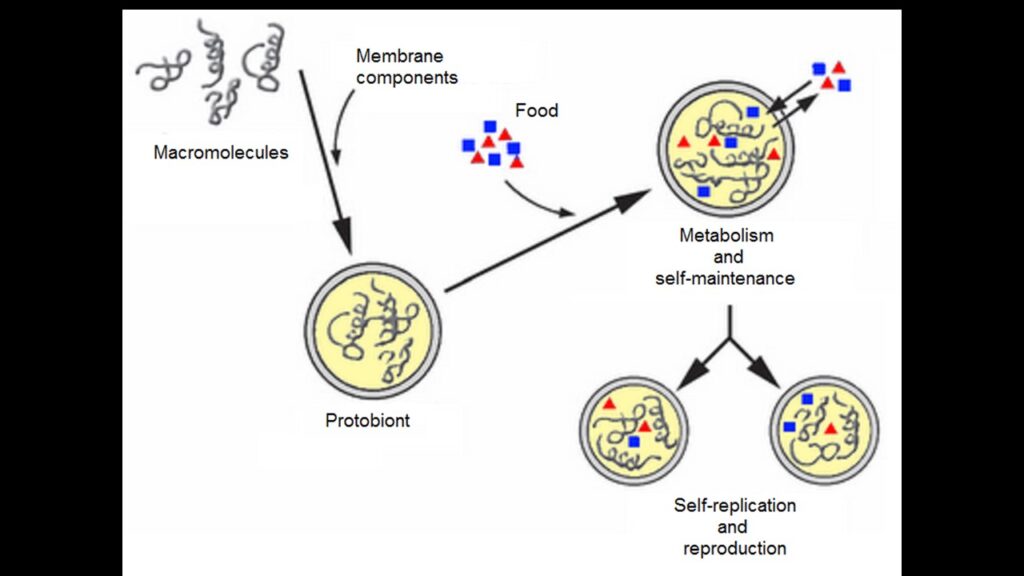
Abb. 3: Vom Replikator zur ersten Zelle
Drittens, und das ist der wichtigste Punkt, gehen die Kreationisten davon aus, dass bei der Abiogenese, wie sie von der Wissenschaft propagiert wird, einige Moleküle zusammenfließen und in einem Augenblick zu einer Zelle werden. Das ist aber nicht der Fall. Wir können nicht einfach ein paar Moleküle wie ein Lego-Set zusammenstecken und eine lebende Zelle erzeugen, denn die Natur hat das auch nicht getan, also hat diese Behauptung nichts mit natürlichen Prozessen zu tun. In Wirklichkeit haben sich Molekülgruppen mit autokatalytischem Verhalten durch natürliche Selektion über lange Zeiträume hinweg entwickelt, um allmählich ein System hervorzubringen, das als lebender Organismus bezeichnet werden kann. Der Zeitpunkt, zu dem ein solches System als Leben bezeichnet werden könnte, wäre in der Tat äußerst schwierig und etwas willkürlich zu definieren. Es gibt ein Gefälle, das so genannte physikalisch-chemische Kontinuum, das chemische Systeme und lebende Organismen miteinander verbindet, und die Grenze zwischen beiden ist unscharf (Abb. 4). Leben und Nichtleben sind kein binäres System wie Kopf und Zahl einer Münze.
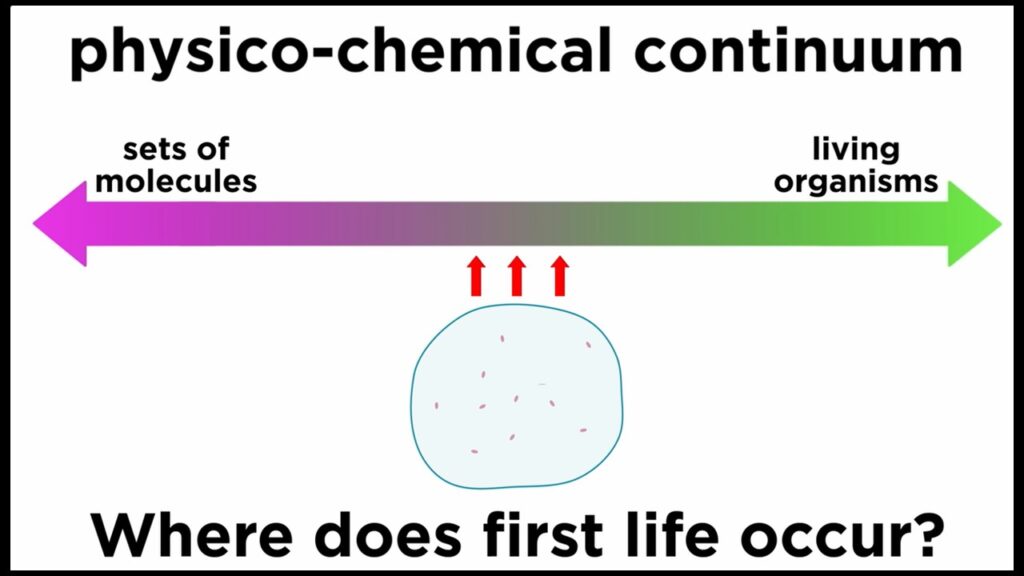
Abb. 4: physikalisch-chemisches Kontinuum
Wenn Kreationisten sich über die Idee lustig machen, dass Leben durch natürliche Prozesse entstanden sein könnte, stellen sie das Produkt der Abiogenese immer falsch dar. Manche betrachten die Komplexität der modernen Tiere, die natürlich völlig verrückt ist, wie bei jeder mehrzelligen Lebensform. Offensichtlich war der erste lebende Organismus einzellig. Deshalb betrachten manche Kreationisten so etwas wie eine Hefezelle. Das ist zutiefst unehrlich, denn Hefezellen sind eukaryotisch und daher das Produkt von Endosymbiose-Ereignissen, die weitaus komplexere einzellige Organismen hervorbrachten, als in den Milliarden Jahren zuvor existierten. Manche gehen noch weiter und untersuchen heutige Bakterienzellen, da diese prokaryotisch sind, aber das ist immer noch völlig unzutreffend. Moderne prokaryotische Organismen sind zwar viel einfacher als einzellige Eukaryonten, aber sie sind immer noch das Produkt von Milliarden von Jahren der Evolution und wesentlich komplexer als die ersten Organismen gewesen sein müssen. Bakterien haben Strukturen wie Flagellen, Zellwände, einfache Organellen und DNA-basierte Genome. Die Diskrepanz in der Komplexität zwischen dem ersten Leben und den heute lebenden Bakterien ist ungefähr so groß wie die zwischen den heute lebenden Bakterien und einem Menschen, vielleicht sogar größer (Abb. 5). Der erste lebende Organismus wäre das gewesen, was wir als Protozelle bezeichnen können. Es wäre die einfachste Struktur gewesen, die die Anforderungen erfüllt, die wir an das Leben stellen, d. h., sie führt einen Stoffwechsel durch und vermehrt sich selbst. Das bedeutet, dass es Enzyme gibt, die in der Lage sind, die Chemie voranzutreiben, und dass sie über ein genetisches Material oder ein Informationsspeichersystem verfügt, von dem die meisten Wissenschaftler glauben, dass es sich um RNA und nicht um DNA handelt.
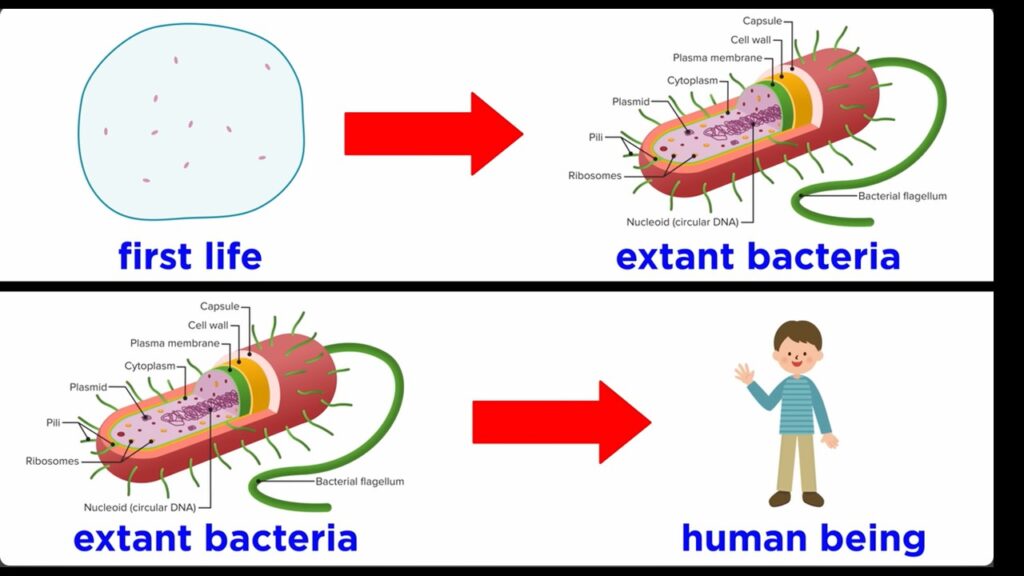
Abb. 5: Unterschied zwischen einem Protobionten und einer Bakterie war in etwa so groß wie zwischen einer Bakterie und einem Menschen.
Die ersten „Lebewesen“ könnten ein einzelnes selbstreplizierendes Molekül gewesen sein, ähnlich dem „selbstreplizierenden“ Peptid der Ghadiri-Gruppe, oder das selbstreplizierende Hexanukleotid, oder möglicherweise eine RNA-Polymerase, die auf sich selbst wirkt (Severin et al. 1997, Lee et al. 1996, 1997a, b, Ekland & Bartel 1996; Abb. 6).
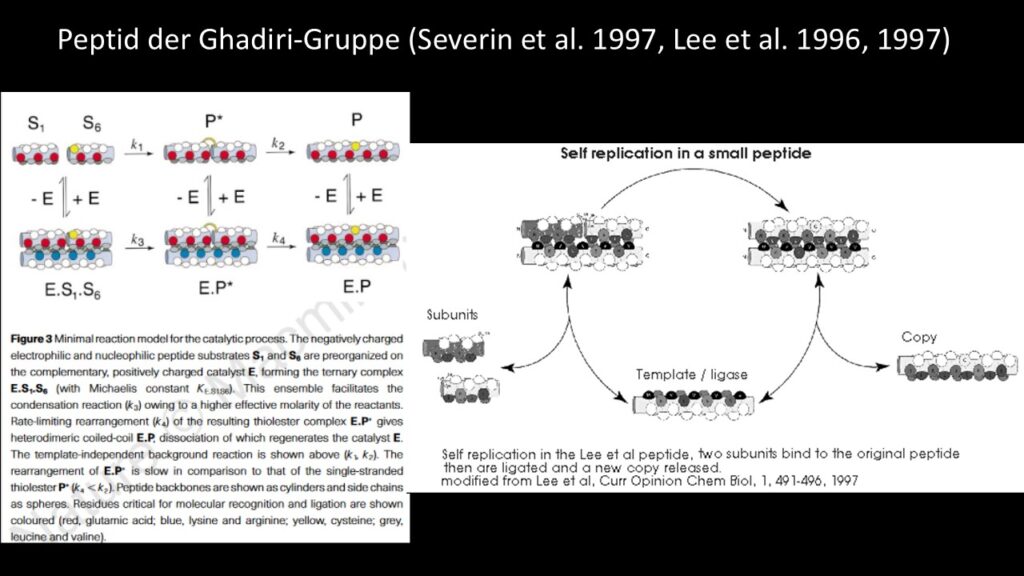
Abb. 6: Peptid der Ghadiri-Gruppe
Nach einer anderen Auffassung waren die ersten Selbstreplikatoren Gruppen von Katalysatoren, entweder Proteinenzyme oder RNA-Ribozyme, die sich in einem katalytischen Zyklus regenerierten (Otsuka & Nozawa 1998, Varetto 1998, Lazcana & Miller 1998, Stadler 1991, Eigen & Schuster 1979). Ein Beispiel ist der SunY-Selbstreplikator mit drei Untereinheiten (Doudna et al. 1991). Diese katalytischen Zyklen könnten in einem kleinen Teich oder einer Lagune begrenzt sein oder ein katalytischer Komplex sein, der entweder an Ton oder an Lipidmaterial auf Ton adsorbiert ist. Oder in einem hydrothermalen Schlot. Da es viele katalytische Sequenzen in einer Gruppe von zufälligen Peptiden oder Polynukleotiden gibt, ist es nicht unwahrscheinlich, dass ein kleiner katalytischer Komplex gebildet werden könnte.
In unseren Beiträgen zur RNA-Welt haben wir den Hyperzyklus von Manfred Eigen und das Chemoton von Ganti kennengelernt (Abb. 7). Im einfachsten Hyperzyklus finden RNA-Moleküle zusammen, die sich in gegenseitiger Wechselwirkung aus einer Substratlösung hervorbringen und „vermehren“. Dabei koppeln sich entweder zwei oder mehrere selbstreproduzierende RNA-Stränge (Ribozyme) oder aber Ribozyme und Enzyme zu einem stabilen Autozyklus, der sich selbst unterhält und repliziert.
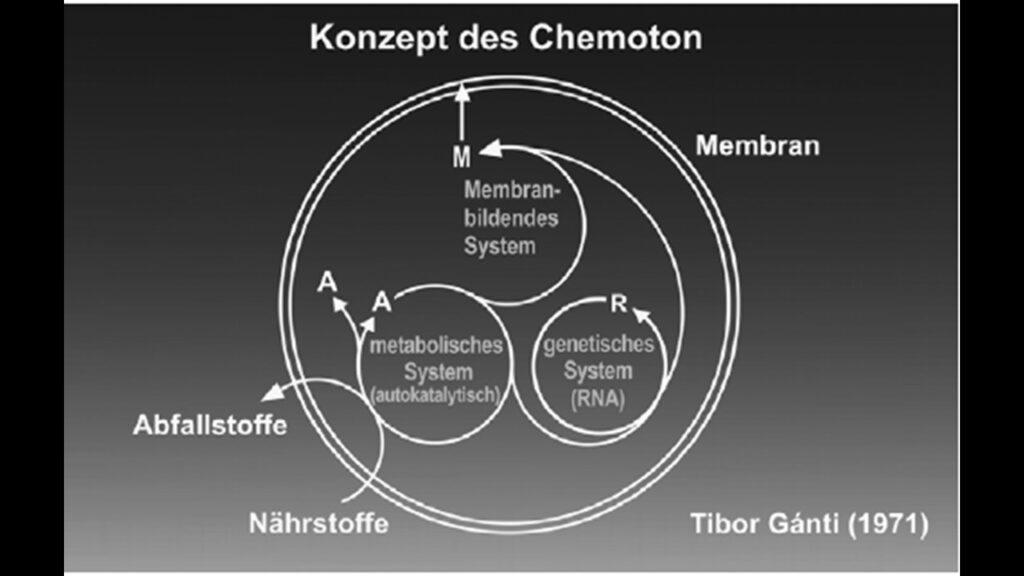
Abb. 7: Chemoton
Unabhängig davon, ob die ersten Selbstreplikatoren einzelne Moleküle oder Komplexe kleiner Moleküle waren, hat dieses Modell nichts mit Hoyles „Tornado auf dem Schrottplatz, der eine 747 baut“ gemein. Um dies zu verdeutlichen, hier ein einfacher Vergleich zwischen der von Kreationisten kritisierten Theorie und der tatsächlichen Theorie der Abiogenese (Abb. 8).
Beachtet, dass die Abiogeneseforschung eine Reihe von kleinen Schritten umfasst – Bildung von Monomeren, dann Bildung von Polymeren, Bildung von Replikatoren, Hyperzyklen etc., und ich habe der Einfachheit halber einige Schritte weggelassen. Bedenkt mit wie viel Zeit und Elan – und an der Fülle an Quellen – ich die Beiträge zur Entstehung des Lebens verfasst habe. Jeder Schritt ist mit einer kleinen Zunahme an Organisation und Komplexität verbunden, und die Chemikalien steigen langsam zum Organismus auf. Die kreationistische Annahme – und so auch ihre Angaben zur Unwahrscheinlichkeit – gegen von einem einzigen großen Sprung aus. Einfache Moleküle und tatdaa schon hat man ein funktionales modernes Protein oder gar eine Zelle. Ich brauche nicht zu wiederholen, dass dies eher das kreationistische Schöpfungsmodell darstellt, als die Abiogenese (Woese 1998, Lee et al. 1997a,b, Lazcarno & Miller 1996, Eigen & Schuster 1979).
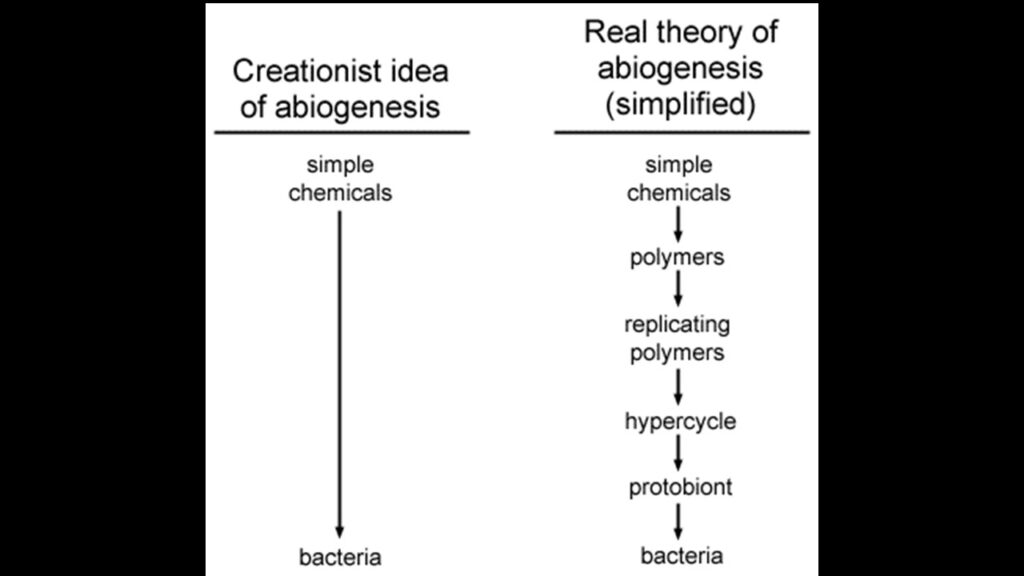
Abb. 8: Unterschied zwischen Abiogenese und Kreationismus
3. Gipfel des Unwahrscheinlichen
Eine Ähnliche Argumentation werfen Kreationisten natürlich auch im Hinblick auf die Rolle der natürlichen Selektion hin. Komplexe Organe oder neue Arten entstünden laut ihnen rein zufällig auf einmal.
Natürliche Selektion ist ein additiver Prozess, der das Problem der Unwahrscheinlichkeit in viele kleine Teile zerlegt. Richard Dawkins hat dies in seinem Buch „Climbing Mount Improbable“ (Gipfel des Unwahrscheinlichen) schön dargestellt (Dawkins 1999). Auf einer Seite des Berges ist eine Felswand, die man unmöglich besteigen kann, aber auf der anderen Seite führt eine sanfte Böschung zum Gipfel. Auf dem Gipfel steht ein komplexes Gebilde, beispielsweise ein Auge oder ein komplexes Protein. Kreationisten haben die absurde Vorstellung, dass solche komplexen Gebilde wie das Auge in einem Schritt entstanden sind, ohne Zwischenstufen. Der Kreationismus symbolisiert also den Sprung vom Fuß der Felswand zum Gipfel. Die Evolution dagegen begibt sich auf die andere Seite des Berges und kriecht über die sanfte Steigung zum Gipfel (vgl. Abb. 9).
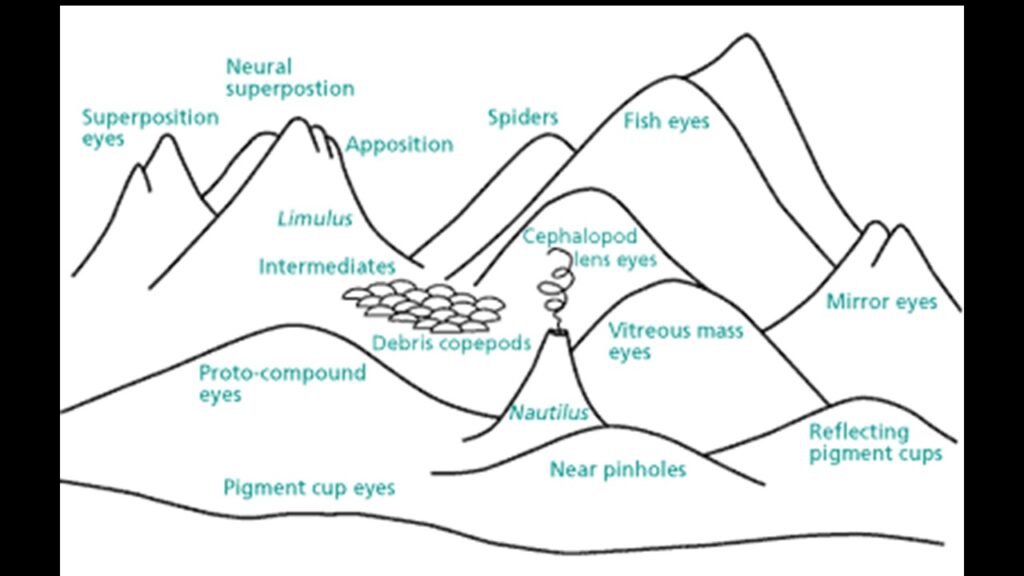
Abb. 9: Gipfel des Unwahrscheinlichen
Hier ist eine Demonstration, wie Mutation und Selektion im Labor funktionieren. Angenommen, wir benötigen ein Enzym, das eine bestimmte Transformation fördert. Wir brauchen eine 100 %ige Ausbeute und eine 100 %ige Spezifität, die hocheffizient durchgeführt wird. Wissenschaftler, die sich in dieser Situation befinden, werden eine gewisse Anzahl von bekannten Enzymen untersuchen, zwischen 100 und 100.000. Die Wahrscheinlichkeit, eine perfekte Übereinstimmung zu finden, ist verschwindend gering. Die Wahrscheinlichkeit, ein Enzym zu finden, das die Umwandlung sehr schlecht durchführt, ist jedoch recht hoch. Vielleicht ergibt das Screening ein Enzym, das die Transformation mit 5 % Ausbeute und 5 % Spezifität durchführt. Man nimmt dieses Enzym und unterzieht es zufälligen Mutationen. Von den Hunderten von Mutanten bietet eine eine Ausbeute von 10 % und eine Spezifität von 20 %. Isolieren wir es und wiederholen den Vorgang. Nach 15 Runden haben wir unser perfektes Enzym. Das Enzym wurde nicht entworfen, es wurde einfach zufällig mutiert und selektiert. Dieser Prozess wird im Labor dramatisch beschleunigt, kann aber in der Natur leicht über längere Zeiträume ablaufen, da ein gewisser Selektionsdruck einfach dadurch entsteht, dass sich bestimmte Molekülsysteme besser selbst reproduzieren als andere. Dies ist der Schlüsselaspekt der Abiogenese, den Kreationisten nicht begreifen können, selbst diejenigen, die über Kenntnisse der traditionellen organischen Chemie verfügen. Die Natur arbeitet mit komplexen Mischungen, sie braucht keine unberührten Bedingungen, wie sie von Laborchemikern verwendet werden. Sie stellt viele Dinge her, und dann übernimmt die Selektion das Ruder, so dass sich bestimmte Sequenzen im Laufe der Zeit durchsetzen. Das Gleiche können wir über lebende Systeme sagen.
Sie sind keine perfekten Maschinen. Organismen haben redundante Funktionen. Sie können nicht wegen Umbau geschlossen werden und müssen Probleme mit Vorhandenem lösen, ohne die Funktionsfähigkeit zu verlieren. Einige Teile haben keine Funktion. Fehlerhafte Gene werden beibehalten. Die Signalwege sind so kompliziert, dass sie von einem technischen Standpunkt aus kaum vorstellbar sind. Ein lebender Organismus ist ein einziges Durcheinander, was im Zusammenhang mit blinden Kräften, die bestehende Vorlagen langsam verbessern, aber nie perfekt machen, durchaus Sinn macht.
4. Der Mythos der „Lebenssequenz“
Eine weitere oft gehörte Behauptung ist, dass es eine „Lebenssequenz“ von 400 Proteinen bzw. Genen gibt und dass die Aminosäuresequenzen dieser Proteine nicht verändert werden können, damit Organismen lebensfähig sind.
Dies ist jedoch Unsinn. Die Behauptung, dass es 400 Proteine gibt, scheint vom proteincodierenden Genom von Mycobacterium genitalium zu stammen (Abb. 10). Sein Erbgut gehört zu den kleinsten bekannten Bakteriengenomen (Mushegian & Koonin 1996). Eine Inspektion des Genoms deutet jedoch darauf hin, dass dieses noch weiter auf einen minimalen Gensatz von 256 Proteinen reduziert werden könnte. Auch hier ist zu beachten, dass es sich um einen modernen Organismus handelt. Der erste Protobiont/Progenote wäre noch kleiner gewesen (Woese 1998), und ihm gingen noch einfachere chemische Systeme voraus (Otsuka & Nozawa 1998, Li et al. 1997a,b, Di Giulio 1997, Lazcano & Miller 1996).
Abb. 10: Proteine von Mycobacterium genitalium
Die Behauptung, dass die Sequenzen von Proteinen nicht verändert werden können, ist ebenfalls Unsinn. In den meisten Proteinen gibt es Bereiche, in denen fast jede Aminosäure ersetzt werden kann, und andere Bereiche, in denen konservative Substitutionen (bei denen geladene Aminosäuren durch andere geladene Aminosäuren, neutrale durch andere neutrale Aminosäuren und hydrophobe Aminosäuren durch andere hydrophobe Aminosäuren ersetzt werden können) vorgenommen werden können. Bei einigen funktionell gleichwertigen Molekülen können zwischen 30 und 50 % der Aminosäuren unterschiedlich sein. Tatsächlich ist es möglich, strukturell nicht identische bakterielle Proteine durch Hefeproteine und Wurmproteine durch menschliche Proteine zu ersetzen, und die Organismen leben recht zufriedenstellend. Die „Lebenssequenz“ ist also ebenfalls ein Mythos.
5. Münzwurf für Anfänger und makromolekularer Aufbau
Spielen wir aber trotzdem das Spiel der Kreationisten und betrachten wir die Bildung eines Peptids durch zufällige Addition von Aminosäuren. Dies ist sicherlich nicht die Art und Weise, wie Peptide auf der frühen Erde entstanden sind, aber es ist lehrreich.
Ich werde als Beispiel das oben erwähnte „selbstreplizierende“ Peptid der Ghadiri-Gruppe verwenden (Severin et al. 1997). Ich könnte auch andere Beispiele verwenden, aber aus Gründen der Einfachheit bleiben wir bei diesem kleinen Peptid.
Dieses Peptid hat eine Länge von 32 Aminosäuren mit der Sequenz RMKQLEEKVYELLSKVACLEYEVARLKKVGE und ist ein Enzym, eine Peptidligase, die aus zwei 16 Aminosäuren langen Untereinheiten eine Kopie ihrer selbst herstellt. Es hat außerdem eine Größe und Zusammensetzung, die ideal für die Bildung durch abiotische Peptidsynthese geeignet ist. Die Tatsache, dass es ein Selbstreplikator ist, ist eine zusätzliche Ironie.
Die Wahrscheinlichkeit, dass diese 32 Aminosäuren lange Sequenz in aufeinanderfolgenden Zufallsversuchen entsteht, beträgt (1/20)32 oder 1 : 4,29 x 1040. Das ist viel, viel wahrscheinlicher als die Wahrscheinlichkeit des vorher erwähnten Beispiels des Proteins aus 300 Aminosäuren, erscheint aber immer noch absurd niedrig.
Es gibt jedoch noch eine andere Seite dieser Wahrscheinlichkeitsschätzungen, und die hängt mit der Tatsache zusammen, dass die meisten von uns kein Gefühl für Stochastik haben. Wenn uns jemand sagt, dass die Wahrscheinlichkeit, dass ein bestimmtes Ereignis eintritt, eins zu einer Million beträgt, erwarten viele von uns, dass eine Million Versuche durchgeführt werden müssen, bevor das besagte Ereignis eintritt, aber das ist falsch.
Hier ist ein Experiment, das Ihr selbst durchführen könnt: Nimmt eine Münze, wirft sie viermal, notiert die Ergebnisse und wiederholt den Versuch. Wie oft, glaubt ihr, müsst ihr diese Prozedur (Versuch) wiederholen, bis ihr viermal hintereinander Kopf erhaltet?
Die Wahrscheinlichkeit für 4 Köpfe in einer Reihe ist (1/2)4 oder 1 : 16: Müssen wir 16 Versuche machen, um 4 Köpfe zu erhalten (KKKK)? Nein, in aufeinander folgenden Experimenten habe ich 11, 10, 6, 16, 1, 5 und 3 Versuche gemacht, um viermal hintereinander Kopf zu haben. Die Zahl 1 zu 16 gibt die Wahrscheinlichkeit eines Ereignisses in einem bestimmten Versuch an, sagt aber nichts darüber aus, wo es in einer Serie auftreten wird. Es kann schon beim ersten Versuch klappen. Selbst bei einer Chance von 1:4,29 x 1040 könnte ein Selbstreplikator überraschend früh auftauchen. Aber da ist noch mehr.
Eine Chance von 1 zu 4,29 x 1040 ist immer noch unglaublich unwahrscheinlich; es ist schwer, mit dieser Zahl umzugehen. Aber, im obigen Beispiel haben wir so getan, als ob nur ein einziges Protein pro Versuch zusammengebaut würde. In Wirklichkeit gäbe es Milliarden von gleichzeitigen Versuchen, wenn die Milliarden von Bausteinmolekülen interagieren (Orgel 1998, Lazcano & Miller 1996).
Kehren wir zu unserem Beispiel mit den Münzen zurück. Nehmen wir an, es dauert eine Minute, die Münzen viermal zu werfen; um viermal Kopf zu haben, bräuchte man im Durchschnitt 8 Minuten. Aber was würde passieren, wenn ihr 16 Freunde darum bittet mit je einer Münze viermal hintereinander Kopf zu werfen, dann beträgt die durchschnittliche Zeit nur eine Minute. Versucht nun, 6-mal hintereinander Kopf zu werfen; die Wahrscheinlichkeit dafür liegt bei (1/2)6 oder 1 zu 64. Das würde im Durchschnitt eine halbe Stunde dauern, aber wenn wir aber 64 Leute rekrutieren, können wir es in einer Minute schaffen. Wenn wir eine Sequenz mit einer Wahrscheinlichkeit von 1:1 Milliarde werfen wollen, rekrutieren wir einfach die Bevölkerung Chinas, um Münzen für uns zu werfen, und wir haben diese Sequenz in kürzester Zeit.
Wenn wir also auf unserer präbiotischen Erde eine Milliarde Peptide gleichzeitig wachsen lassen, verkürzt sich die Zeit für die Erzeugung unseres Replikators erheblich.
Und obwohl eine Milliarde Ausgangsmoleküle eine Menge Moleküle sind, könnten wir jemals genug Moleküle bekommen, um unseren ersten Replikator in weniger als einer halben Milliarde Jahre zufällig zusammenzusetzen?
Ja, ein Kilogramm der Aminosäure Arginin enthält 2,85 x 1024 Moleküle (das sind weit über eine Milliarde Milliarden); eine Tonne Arginin enthält 2,85 x 1027 Moleküle. Wenn man eine Sattelschlepperladung jeder Aminosäure in einen mittelgroßen See kippen würde, hätte man genug Moleküle, um unseren speziellen Replikator in ein paar Dutzend Jahren zu erzeugen, wenn man bedenkt, dass man 55 Aminosäuren lange Proteine in 1 bis 2 Wochen herstellen kann (Ferris et al. 1996, Ertem & Ferris 1996).
Wie verhält sich das nun mit der präbiotischen Erde? Auf der frühen Erde hatte der Ozean wahrscheinlich ein Volumen von 1 x 1024 Litern. Bei einer Aminosäurekonzentration von 1 x 10-6 M (eine mäßig verdünnte Suppe, siehe Chyba & Sagan 1992 [23]) gibt es etwa 1 x 1050 potenzielle Ausgangsketten, so dass eine beträchtliche Anzahl effizienter Peptidligasen (etwa 1 x 1031) in weniger als einem Jahr, geschweige denn in einer Million Jahren hergestellt werden könnte. Die Synthese primitiver Selbstreplikatoren könnte relativ schnell vonstattengehen, selbst bei einer Wahrscheinlichkeit von 1:4,29 x 1040.
Nehmen wir an, dass es eine Woche dauert, eine Sequenz zu erzeugen (Ferris et al. 1996, Ertem & Ferris 1996). Dann könnte die Ghadiri-Ligase in einer Woche erzeugt werden, und jede beliebige Cytochrom-C-Sequenz oder andere komplexe Enzyme könnten in etwas mehr als einer Million Jahren erzeugt werden.
Ich möchte nochmals anmerken: ich sage nicht, dass die ersten Proteine und das Leben genauso auf der Urerde entstanden sind – wie das passierte, das habe ich in den vorherigen Beiträgen gezeigt. Ich habe lediglich das „Wahrscheinlichkeitsargument“ der Kreationisten für bare Münze genommen.
Ich habe also gezeigt, dass die Erzeugung eines bestimmten kleinen Proteins nicht so verblüffend schwierig ist, wie Kreationisten annehmen – selbst wenn man ihre hirnrissigen Annahmen zur Entstehung komplexer Proteine in einem Schritt ernst nimmt.
6. Laborbedingungen
Kreationisten behaupten erstens, dass jede Laborsynthese nicht auf die Abiogenese anwendbar ist. Sie geben vor, Aspekte des Versuchsaufbaus und der verwendeten Reagenzien in Frage zu stellen, und behaupten, dass die Reagenzien auf einer präbiotischen Erde nicht verfügbar waren, die Ausrüstung nicht zur Verfügung stand, die Manipulationen am Kolben nicht stattgefunden hätten und so weiter. Im Grunde genommen zählt alles, was in irgendeinem Labor gemacht wurde, nicht. Das ist höchst unredlich. Im Labor können wir testen, was Moleküle tun. Wenn sie in einem Kolben etwas tun, dann ist das durch die Eigenschaften dieser Moleküle bedingt. Sie werden überall die gleiche Chemie betreiben. Wenn wir den Siedepunkt einer Verbindung messen, dann ist das nicht nur in einem Labor möglich. Es ist die Temperatur, bei der diese Verbindung bei einem bestimmten Druck siedet, und zwar überall im Universum. Das Labor ist keine Zauberei, und die Natur ist es auch nicht. Wir können Moleküle nicht dazu zwingen, in einem Labor Dinge zu tun, die sie in der Natur unter den gleichen Bedingungen nicht tun. Außerdem wird bei Experimenten meist nur eine Sache auf einmal getestet. Es wird ein Aspekt der präbiotischen Chemie untersucht, so dass tangentiale Prozesse mit Reagenzien, die nicht präbiotisch sind, für die Forschungsfrage nicht relevant sind. Kreationisten fixieren sich auf irrelevante Details, um vom Sinn eines Experiments abzulenken. Das Gleiche tun sie mit dem Versuchsaufbau. Wenn irgendeine Art von Maschinerie verwendet wird, schreien sie auf. Auch das ist kurzsichtig, denn es gibt bestimmte geochemische oder geophysikalische Prozesse und Zyklen in der Natur, die im Labor simuliert werden müssen. Der Aufbau ahmt gezielt die natürlichen Bedingungen nach. Dies gilt auch für bestimmte Aspekte des Versuchsprotokolls, wie die Durchführung von Reaktionen unter Argongas oder das Waschen von Mineraloberflächen mit Peroxiden und entionisiertem Wasser. Die Reaktionen werden unter Argongas durchgeführt, um zu verhindern, dass das Reaktionsgefäß mit der Atmosphäre interagiert, d. h. mit all dem Sauerstoff, der auf der präbiotischen Erde nicht vorhanden war. Das Waschen mit Peroxiden und entionisiertem Wasser wird durchgeführt, um biologische Verunreinigungen wie Bakterien abzutöten und zu entfernen, die auf der präbiotischen Erde offensichtlich nicht vorhanden waren. Diese Art von Protokoll macht die Experimente nicht weniger, sondern mehr präbiotisch relevant. Forscher, die sich mit dem Ursprung des Lebens befassen, sind weder dumm, noch sind sie betrügerisch. Sie gehen die Dinge aus bestimmten Gründen auf eine bestimmte Art und Weise an, die Kreationisten, die nicht das Geringste von der Durchführung chemischer Reaktionen verstehen, völlig unbekannt sind. Und natürlich gibt es Aspekte der Laborchemie, die ausschließlich zu dem Zweck durchgeführt werden, dass der Forscher feststellen kann, was passiert ist. Wenn Chemiker ihre Produkte isolieren und reinigen, dann nur, um sie mit Hilfe spektroskopischer Techniken zu charakterisieren, die es ihnen ermöglichen, herauszufinden, was genau sie erhalten haben und ob es das ist, was sie erwartet haben. Wir können nicht erkennen, was in einem Kolben ist, wenn wir ihn nur mit den Augen ansehen. Wir brauchen spezielle Instrumente, und die Verbindungen müssen für die Analyse rein sein. Die Natur muss das nicht tun, denn sie hat keine Gefühle und kümmert sich nicht darum, was passiert. Sie stellt Dinge her und macht einfach weiter. Die andere Sache, die Kreationisten tun, ist, das Konzept der „Zeit der Lücken“ zu erfinden, die Vorstellung, dass Wissenschaftler annehmen, dass mit genügend Zeit alle relevanten chemischen Vorgänge stattfinden werden. Dies ist ein offensichtlicher Versuch, die Aufmerksamkeit von ihrer eindeutigen Gott-der-Lücken-Argumentation abzulenken, aber darüber hinaus ist es nur eine weitere Täuschung ihrerseits. Die Behauptung lautet, dass sich Moleküle sehr schnell zersetzen, so dass Prozesse, die lange Zeiträume erfordern, unmöglich werden, weil die Reaktionspartner nicht intakt bleiben. Bei diesen Argumenten werden die Abbauraten immer maßlos übertrieben oder falsch dargestellt, entweder aus Unwissenheit oder durch Taschenspielertricks, indem die Halbwertszeit einer einzelnen Peptidbindung unter bestimmten Bedingungen auf ein ganzes Peptid unter beliebigen Bedingungen extrapoliert wird. Sie ignorieren auch die stabilisierende Wirkung mineralischer Träger, die den Abbau vermindern und gleichzeitig die Polymerisation fördern. Und das größte Versäumnis ist natürlich, dass es gut ist, dass Polymere degenerieren, weil dadurch ein Gleichgewicht zwischen Monomeren und Polymeren hergestellt wird, innerhalb dessen sich für verschiedene Polymere unterschiedliche Geschwindigkeitskonstanten ergeben, wodurch eine Art natürliche Auslese stattfindet, bei der sich einige Polymere schneller bilden oder langsamer abbauen oder effektiver stabilisiert werden, und dies sind Faktoren, die dazu führen, dass bestimmte Sequenzen in statistisch signifikanter Weise überwiegen.
7. Thermodynamik
Das nächste Thema, das von Kreationisten im Zusammenhang mit der Entstehung von Leben totargumentiert wird, ist der zweite Hauptsatz der Thermodynamik und die Entropie. Entropie ist, grob gesagt, ein Maß für die Verteilung von Materie und Energie in einem System (Abb. 11). Wenn also die Moleküle hoch konzentriert und organisiert sind, handelt es sich um ein System mit geringer Entropie, sind die Moleküle dagegen weit verstreut und ungeordnet, handelt es sich um ein System mit hoher Entropie. Was den zweiten Hauptsatz der Thermodynamik betrifft, so behaupten einige Kreationisten, dass dieses Gesetz die spontane Bildung von Ordnung verbietet, was nicht der Fall ist, und sie wiederholen diese Unwahrheit bei jeder Gelegenheit. Offensichtlich bilden sich geordnete Strukturen immer wieder spontan. Wassermoleküle gefrieren und bilden schön gemusterte Eiskristalle. Es bilden sich hochgradig geordnete Mineralien und Seifenmoleküle aggregieren spontan zu Mizellen, ein Prozess, der auch die Bildung der ersten Vesikelmembranen aus analogen Molekülen wie einfachen Fettsäuren erklärt.
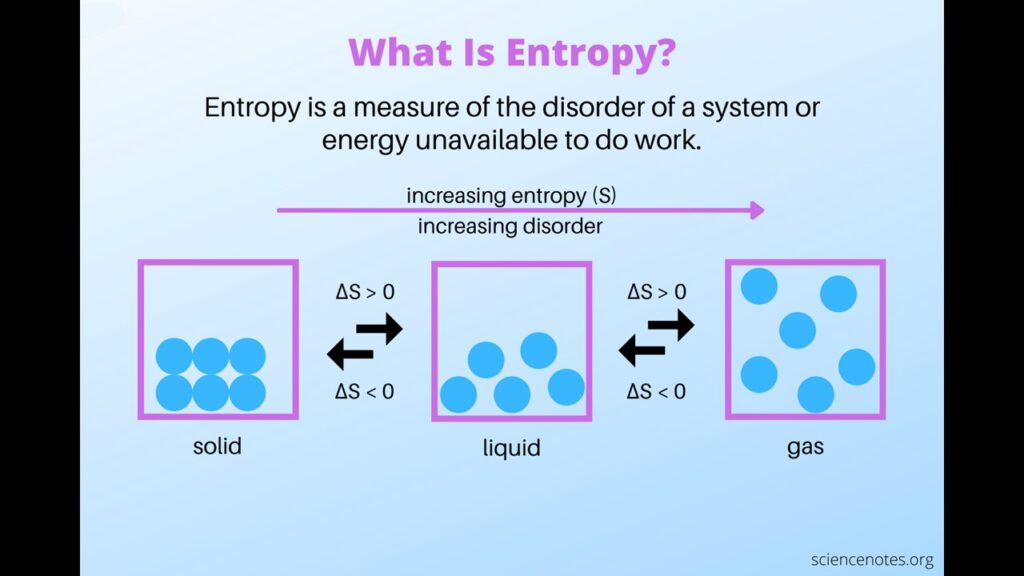
Abb. 11: Entropie
Lokalisierte geordnete Strukturen sind nicht nur möglich, sondern werden von der Thermodynamik gezielt gefördert. Dieses Gesetz besagt nämlich, dass die Entropie des Universums bei jedem spontanen Prozess zunehmen muss. Die Spontaneität wird durch die Änderung eines Parameters bestimmt, der als freie Gibbs-Energie bezeichnet wird, dargestellt durch delta G. Diese wiederum wird durch die Änderung der Enthalpie, delta H, bestimmt, die im Wesentlichen als Wärmeaustausch bei konstantem Druck angesehen werden kann, sowie durch die Änderung der Entropie, delta S, und auch der Temperatur, T, gemäß der folgenden Gleichung (abb. 12).

Abb. 12: Entropie und Gibbsenergie
Das erste Problem liegt in der Annahme, dass dieses Gesetz auf Systeme wie einen lebenden Organismus oder sogar die Erde selbst zutrifft. Bei beiden handelt es sich um offene Systeme ohne Gleichgewicht, die ständig in Bewegung sind und sowohl Energie als auch Materie mit ihrer Umgebung austauschen (Abb. 13). Wenn lokalisierte geordnete Strukturen entstehen, wie z. B. Wassermoleküle, die Eis bilden, führen die neuen Wechselwirkungen zwischen den Wassermolekülen, die so genannten Wasserstoffbrückenbindungen, dazu, dass das System aus Molekülen Energie freisetzt. Das System der Moleküle geht also auf eine niedrigere Energie über, und diese Energie wird an die Umgebung abgegeben. Dies führt zu einer Ausbreitung der Energie und somit zu einer Zunahme der Entropie im Universum. Zoomt man näher heran, so hat die Entropie lokal abgenommen, da die Wassermoleküle geordneter werden, d. h. Delta S ist negativ, aber sie haben Energie freigesetzt, d. h. Delta H ist negativ, was zu einem negativen Wert für Delta G führt, so dass der Prozess in Bezug auf das System der Moleküle spontan ist. Zoomt man wieder heraus, so bedeutet die Freisetzung von Energie eine Zunahme der Entropie für das System und die Umgebung, auch bekannt als das Universum. Die lokale Abnahme der Entropie ist irrelevant, es ist die Zunahme der Entropie für das Universum, die durch den zweiten Hauptsatz der Thermodynamik festgelegt ist. Dies gilt auch für die Polymerisation und Aggregation von Molekülen, die vollkommen spontan ablaufen.
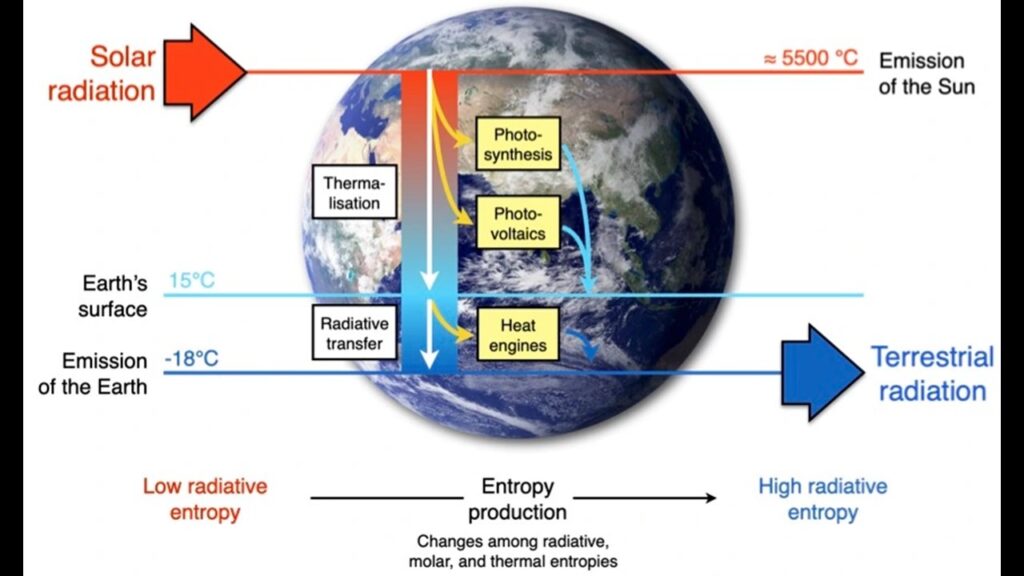
Abb. 13: Die Erde als offenes System
Kreationisten, die behaupten, dass die Thermodynamik die Selbstorganisation geordneter Systeme nicht zulässt, haben also absolut keine Ahnung, wovon sie reden, oder sie lügen, oder beides. Um noch weiter zu gehen: Seit Jahrzehnten werden zahlreiche Studien darüber durchgeführt, wie Systeme, die einen konstanten Energiefluss erhalten, wie die Erde, die konstantes Sonnenlicht erhält, unweigerlich geordnete Strukturen erzeugen, um diese externen Gradienten freier Energie zu zerstreuen, um sich von einer Gleichgewichtsposition wegzubewegen und Energie freizusetzen, was zu einem Anstieg der Entropie im Universum führt. Im Gegensatz zu den Behauptungen der Kreationisten kann man mit Fug und Recht behaupten, dass bei solchen Nicht-Gleichgewichtssystemen die Entropie die eigentliche Triebfeder für die Bildung von lokal geordneten Strukturen ist, von Wirbelstürmen über Galaxien bis hin zu lebenden Organismen. Deutlicher kann man es nicht machen. Es ist insbesondere der zweite Hauptsatz der Thermodynamik, der zu einer Zunahme der Ordnung führt und letztlich Leben unvermeidlich macht, wenn genügend Zeit und Ressourcen zur Verfügung stehen.
8. Information
Ein weiterer Irrtum der Kreationisten hat mit dem Begriff der Information zu tun. Es ist richtig, dass ein RNA-Strang, der sich selbst repliziert, Informationen enthält und überträgt. Tatsächlich enthält jeder RNA-Strang Informationen, unabhängig davon, ob er funktional ist oder nicht. Die Information ist die Sequenz der Nukleotide im Strang, die aus einer Kombination von A, U, G und C besteht. So wie Wörter aufgrund ihrer Buchstaben Informationen enthalten, gilt das auch für diese Biopolymere mit unterschiedlichen Untereinheiten. Das Wichtigste ist die funktionelle Information, die die Wahrscheinlichkeit darstellt, dass eine Konfiguration eine bestimmte Funktion erfüllt. Kreationisten werden daher unablässig fragen: Woher kommt die Information? Manche bezeichnen sie sogar als „spezifizierte Information“, was bedeutet, dass sie von einem Schöpfer spezifiziert worden sein muss, da sie eine so präzise Funktionalität ergibt. Andere Kreationisten behaupten, die Information sei eine Art Essenz oder materielle Substanz, die das Molekül neben der Sequenz selbst besitze und deren Ursprung erklärt werden müsse. Wenn Ribonukleotide polymerisieren, entsteht eine Sequenz. Das ist die Information. Jede Sequenz von Nukleotiden besitzt Information, so dass der Prozess der Nukleotidpolymerisation notwendigerweise Information hervorbringt. Es ist der Prozess der Selektion, der auf der Replikationsrate und -treue beruht, der diese Information sinnvoll macht. Wer also fragt, „woher die Information kommt“, hat weder verstanden, was Information ist, noch was Biopolymere sind.
Zusammenfassend lässt sich sagen, dass Kreationisten das Thema Abiogenese von allen Themen, die sie zu erörtern versuchen, am schlechtesten abhandeln. Das Ausmaß der Verzerrung, das bei der Gleichsetzung der Komplexität des existierenden Lebens mit dem, was die erste Urzelle gewesen sein muss, oder den Mechanismen, durch die sich ihre Bestandteile gebildet haben können, auftritt, wird selbst von den schrägsten Einschätzungen der Evolution nicht übertroffen, weil diese Konzepte so kompliziert sind. Auch wenn dies sicherlich eine stark verkürzte Version dessen war, was wir über den Ursprung des Lebens wissen, so dient es doch hoffentlich dazu, die tiefgreifende Kurzsichtigkeit zu verdeutlichen, mit der Kreationisten an dieses komplizierte Thema herangehen. Die Abiogenese ist ein unheimlich komplexes Gebiet, das viele Teildisziplinen miteinschließt. Wie mannigfaltig die Forschung ist, zeigt meine eigene Videoreihe zum Thema der Entstehung des Lebens, welche die hier angesprochenen Aspekte in dutzenden von halbstündigen Videos beschreibt, in der auch Aspekte angesprochen werden, die jenseits der hier angesprochenen Themen liegen, wie dem Ursprung des genetischen Codes oder der Stoffwechselprozesse.
Literatur
Chyba C, Sagan C (1992): Endogenous production, exogenous delivery and impact-shock synthesis of organic molecules: an inventory for the origins of life. Nature, 355: 125-32.
Dawkins, R (1999): Gipfel des Unwahrscheinlichen. Reinbek: Rowohlt
Di Giulio M (1997): On the RNA world: evidence in favor of an early ribonucleopeptide world. J Mol Evol, 45: 571-8
Doudna JA, Couture S, Szostak JW (1991): A multisubunit ribozyme that is a catalyst of and template for complementary strand RNA synthesis. Science, 251: 1605-8
Eigen M, Schuster P, (1979): The hypercycle. A principle of natural self-organization. Springer-Verlag, isbn 3-540-09293
Ekland EH, Bartel DP (1996): RNA-catalysed RNA polymerization using nucleoside triphosphates. Nature, 383: 192
Ertem G, Ferris JP (1996): Synthesis of RNA oligomers on heterogeneous templates. Nature, 379: 238-40
Ferris JP, Hill AR Jr, Liu R, Orgel LE (1996): Synthesis of long prebiotic oligomers on mineral surfaces [see comments]. Nature, 381: 59-61
Lazcano A, Miller SL, (1996): The origin and early evolution of life: prebiotic chemistry, the pre- RNA world, and time. Cell, 85: 793-8
Lee DH, Granja JR, Martinez JA, Severin K, Ghadri MR (1996): A self-replicating peptide. Nature, 382: 525-8
Lee DH, Severin K, Ghadiri MR. (1997a): Autocatalytic networks: the transition from molecular self-replication to molecular ecosystems. Curr Opinion Chem Biol, 1, 491-496
Lee DH, Severin K, Yokobayashi Y, Ghadiri MR, (1997b): Emergence of symbiosis in peptide self-replication through a hypercyclic network. Nature, 390: 591-4
Mushegian AR, Koonin, EV (1996): A minimal gene set for cellular life derived by comparison of complete bacterial genomes. Proc. Natl. Acad. Sci. USA, 93: 10268-10273.
Orgel LE (1998): Polymerization on the rocks: theoretical introduction. Orig Life Evol Biosph, 28: 227-34
Otsuka J, Nozawa Y (1998): Self-reproducing system can behave as Maxwell’s demon: theoretical illustration under prebiotic conditions. J Theor Biol, 194, 205-221
Ruse M, (1997): The origin of life, philosophical perspectives. J Theor Biol, 187: 473-482
Severin K, Lee DH, Kennan AJ, Ghadiri MR (1997): A synthetic peptide ligase. Nature, 389: 706-9
Stadler PF (1991): Dynamics of autocatalytic reaction networks. IV: Inhomogeneous replicator networks. Biosystems, 26: 1-19
Varetto L, (1998): Studying artificial life with a molecular automaton. J Theor Biol, 193: 257-85
Woese C, (1998): The universal ancestor. Proc Natl Acad Sci USA, 95: 6854-6859.
[1] https://www.talkorigins.org/faqs/abioprob/abioprob.html
[2] https://www.cambridge.org/core/books/failures-of-mathematical-antievolutionism/A95C44708846F7439E7997D5832E2241
