
Die Frage, warum es eben nur zwei Geschlechter geben kann, hat mit ihrer biologischen Funktion zu tun. Hierbei geht es um Sex, Chromosomenzahl und Keimzellenbildung.
Sexualität, bzw. sexuelle Fortpflanzung erschöpft sich nicht nur in der Vermehrung der Individuenzahl, sondern beruht auf der ständigen Neukombination des elterlichen Erbgutes. Damit ist eine gewisse Variabilität der Organismen gesichert. Sex bezeichnet die Gesamtheit aller Phänomene, die in diesen Dienst gestellt werden. Über die Ursprünge und Bedeutung der Sexualität hat schon der Biologe August Weismann hingewiesen (WEISMANN 1883, 1886). Über die Ursprünge und Evolution der sexuellen Fortpflanzung hat LANE (2009) ein Kapitel gewidmet; MARGULIS & SAGAN (1986) haben hierzu ebenfalls ein wunderbares Buch geschrieben (wenn auch teilweise betagt). Für visuell orientierte Menschen empfehle ich folgendes Video, außerdem noch folgendes
Chromosomen, haploid oder diploid?
Wie wird diese Variabilität erzeugt? Sexualität ist die Verschmelzung zweier Zellen und ihrer Genome. Hierfür werden Keimzellen produziert. Wichtig ist hier anzumerken, dass der Großteil der sich sexuell fortpflanzenden Organismen über einen doppelten Chromosomensatz verfügt. Chromosomen sind Bestandteile der Zelle, auf denen die Erbinformationen gespeichert sind. Sie bestehen jedoch nicht nur aus der DNA, sondern auch aus einer Reihe von Proteinen, zumeist sogenannte Histone (Abb. 1). Der Komplex aus DNA und Proteinen wird Chromatin genannt. Die DNA liegt also nicht frei im Zellkern, sondern liegt in einer verdichteten (kondensierten) Form vor. Bei der Zellteilung (Mitose) hat das Chromatin als sogenannte Metaphase-Chromosomen (Metaphase bezeichnet ein Stadium der Mitose) die dichteste Kondensation (vgl. ALBERTS et al. 2017, CZIHAK 1981, WOLPERT et al. 2007).
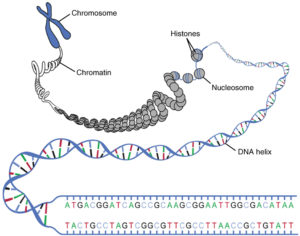
Abb. 1: Sogenannte Kondensation der DNA. Die DNA findet sich als Doppelstrang, der eine Helix bildet. Dieser DNA-Faden windet sich um die Histone. 4 Histone bilden zusammen ein Nukleosom. Die Nukleosome winden sich ebenfalls und bilden das Chromatin-Gerüst. Die Gesamtheit bezeichnet man als Chromosom. Quelle: Lecturio
Jede Tierart verfügt über eine bestimmte Anzahl an Chromosomen. Wir Menschen haben 46 Chromosomen. 44 sogenannte autosomale Chromosomen und 2 Geschlechtschromosomen (Frauen: XX, Männer: XY). Tatsächlich handelt es sich bei den 46 Chromosomen, die in 23 Paaren angeordnet sind. Das heißt: Wir haben 23 unterschiedliche Chromosomen, die jeweils doppelt vorhanden sind. Doppelte Chromosomen sind zueinander homolog (sie sind Variationen desselben Chromosomentyps). Einen doppelten Chromosomensatz bezeichnet man als diploid oder kurz: 2n. Chromosomensätze können in sogenannten Karyogrammen dargestellt werden (siehe Abb. 2, vgl. ALBERTS et al. 2017, CZIHAK 1981, WOLPERT et al. 2007).
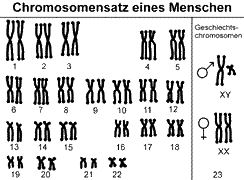
Abb. 2 Karyogramm eines Menschen (Metaphasenchromosomen, bei der das Erbgut schon verdoppelt ist und in der Endphase der Mitose auf die zwei Tochterzellen verteilt wird). Quelle: Digitalfolien
Nerd-Wissen: Unsere nächsten Verwandten, die Schimpansen & Bonobos, teilen zwar 98-99% unseres Erbgutes, doch haben sie statt 46 Chromosomen 48; d. h. sie haben ein Chromosomenpaar mehr. Für Kreationisten ist das übrigens der „Beweis“, dass wir unmöglich mit den Affen verwandt sein können. Offensichtlich vergessen sie, dass eine Arbeitsgruppe des „Chimanzee genome project“ 2002 nachweisen konnte, dass unser Chromosom Nr. 2 verschmolzen ist und bei Schimpansen dieser in zwei Chromosomen aufgeteilt ist (2A & 2B). Als vor 6 Mio. Jahren sich die Evolutionswege von Mensch und Schimpanse trennten, hatte unsere Evolutionslinie die beiden Chromosomen 2A und 2B zu einen Chromosom verschmolzen. So konnten alle Gene erhalten bleiben. Chromosom 2 des Menschen ähnelt Chromosom 2A und 2B des Schimpansen so sehr, dass es sich um eine Fusion dieser handeln musste. Am Ort der Fusion findet sich eine etwa 150.000 Basenpaar lange Stelle, die sich bei den Schimpansen nicht findet und für 4 Gene codiert. Kopien dieser Gene finden sich auch im menschlichen Chromosom Nr. 9. (vgl. Abb. 3; DEGROUCHY 1987, FAN et al. 2002 ).
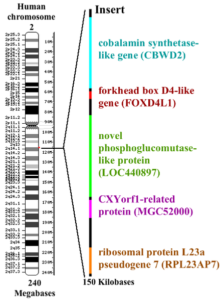
Abb. 3: Fusion der Schimpansen-Chromosomen 2A & 2B zu Human-Chromosom Nr. 2. Der Rote Punkt zeigt die Fusionsstelle beider Chromosomen, an der eine 150.000 Basenpaar lange DNA-Sequenz inseriert (rechts vergrößert dargestellt). Quelle: wikipedia
Der doppelte Chromosomensatz kommt dadurch zustande, dass wir die eine Hälfte von unserem Vater, die andere von unserer Mutter erhalten haben. Es haben sich also die Chromosomen von zwei verschiedenen Zellen verschmolzen und bildeten so unseren doppelten Chromosomensatz. Unsere Keimzellen, welche miteinander verschmelzen sind haploid (kur 1n), d. h. sie haben nur einen Chromosomensatz. Der Prozess, bei dem haploide Zellen entstehen, bezeichnet man als Meiose, die von der eigentlichen Zellteilung (Mitose) unterschieden wird (vgl. ALBERTS et al. 2017, CZIHAK 1981, WOLPERT et al. 2007).
Mitose – Zellteilung
Die Zellteilung (Mitose, Abb. 4, vgl. ALBERTS et al. 2017, CZIHAK 1981, WOLPERT et al. 2007) findet in verschiedenen Schritten statt.
Die Interphase ist der Prozess zwischen zwei Zellteilungen. In der G1-Phase – manchmal auf Gap-Phase genannt – wächst die Zelle sehr stark und bereitet sich auf die Zellteilung vor. Dabei findet eine Steigerung der Proteinsynthese statt. In der S-Phase findet die Verdoppelung (Replikation) der DNA statt. Die G2-Phase stellt den Übergang zur Mitose dar; alle Voraussetzungen für die Kernteilung sind nun vorhanden.
In der Prophase ziehen sich die Chromosomen stark zusammen. Dabei kondensieren sie und werden durch diesen Vorgang unter dem Lichtmikroskop sichtbar. Jedes Chromosom besteht aus zwei identischen Strängen (Chromatiden genannt = Zwei-Chromatid-Chromosomen; nicht zu verwechseln mit Chromatin!), die sich größtenteils voneinander trennen und nur noch durch einen so genannten Centromer zusammengehalten werden. Parallel beginnt die Ausbildung der Mitose-Spindel, einem Teil des Cytoskeletts, welches die Aufgabe hat, die einzelnen Chromatiden voneinander zu trennen.
In der Metaphase haben sich die Chromosomen (bestehend aus je zwei Chromatiden= Zwei-Chromatid-Chromosomen) noch weiter verkürzt (stärkste kondensierte Form) und in einer Ebene angeordnet (Äquatorialebene). Die Schwesterchromatiden weisen zu den entgegengesetzten Spindelpolen. Diese bilden Spindelfäden, die sich an die Centromere der Chromosomen heften.
In der Anaphase verkürzen sich die Spindelfäden und trennen die einzelnen Chromatiden voneinander. In der Telophase entstehen so zwei diploide Zellen (2n), also beim Menschen mit 46 Chromosomen (=Ein-Chromatid-Chromosomen).
So wird versichert, dass die Zellen bei der Teilung ihren jeweiligen Chromosomensatz erhalten.
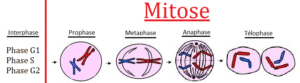
Abb. 4. Mitose. Quelle: https://jeretiens.net/difference-entre-mitose-et-meiose/
Meiose – Reduktionsteilung
Die Meiose, also die Bildung haploider (1n) Keimzellen, verläuft ähnlich ab (Abb. 5, vgl. ALBERTS et al. 2017, CZIHAK 1981, WOLPERT et al. 2007). Die Meiose unterteilt man zur besseren Übersicht in verschiedenen Phasen, bei der man grob gesagt eine erste Reifeteilung und eine zweite Reifeteilung unterscheidet. Auch hier wird während der Interphase die DNA verdoppelt, man erhält also Chromosomen mit je zwei Chromatiden, die in der Prophase 1 kondensieren.Die Chromosomen paaren sich so, dass beide Kopien von Chromosom 1 zusammen sind, beide Kopien von Chromosom 2 zusammen sind und so weiter.
In der Metaphase 1 der Meiose ordnen sich nun die homologen Chromosomen beiderseits der Äquatorialebene an und je ein Chromosom (bestehend aus zwei Chromatiden) weist zu einem Spindelpol. Hier ist der Unterschied zur Metaphase der Mitose zu sehen, bei dem sich nicht die homologen Chromosomenpaare anordnen, sondern jedes einzelne Chromosom.
Während der Anaphase 1 werden die einzelnen Chromosomen an die Spindelpole gezogen, so entstehen während der Telophase 1 zwei Zellen mit dem halben Chromosomensatz (1n). Es handelt sich hierbei aber immer noch um Zwei-Chromatid-Chromosomen, d. h. die DNA liegt hier immer noch doppelt vor.In der Meiose I bleiben die Schwesterchromatiden zusammen. Dies ist anders als bei Mitose und Meiose II. Folgerichtig kommt es zur zweiten Reifeteilung, die genauso abläuft wie die Mitose. Am Ende entstehen 4 haploide Zellen (1n) mit Ein-Chromatid-Chromosomen.
Weil bei der Meiose die Chromosomenzahl reduziert wird, spricht man auch von einer Reduktionsteilung.
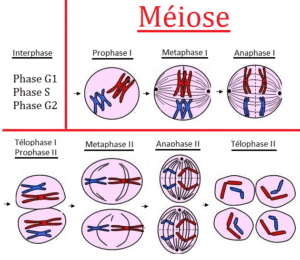
Abb. 5: Meiose. Quelle: https://jeretiens.net/difference-entre-mitose-et-meiose/
Der Meiose-Prozess klingt erstmal kompliziert, doch der wesentliche Aspekt, den wir hier festhalten können, ist die Tatsache, dass bei der Meiose haploide Zellen entstehen (1n). Diese haploiden Zellen bezeichnet man auch als Keimzellen, bzw. Gameten. Eine Verschmelzung (=Befruchtung) zweier Gameten lässt wieder eine diploide (2n) Zelle (die Zygote) entstehen, aus der sich durch mitotische Teilungen ein vielzelliger Organismus entwickelt.
Hier haben wir schon das erste Argument, weshalb es biologisch betrachtet nur zwei Geschlechter geben kann: Da wir über einen doppelten Chromosomensatz verfügen, sind wir das Produkt der Verschmelzung von ZWEI haploiden Keimzellen (Gameten).
Kernphasenwechsel
Organsimen haben dabei also immer einen Wechsel zwischen einer haploiden Phase (Gameten) und einer diploiden. Dieser regelmäßige Wechsel zwischen einem durch die Meiose eingeleiteten haploiden Zustand (Haplophase) und einem aus der Befruchtung resultierenden diploiden Zustand (Diplophase) ist für alle Eukaryoten (Zellen, die über einen Zellkern verfügen. Dazu zählen alle Tiere, Pflanzen, Pilze, Algen und tierischen Einzeller), die sich sexuell Vermehren, kennzeichnend. Je nachdem welche der beiden Phasen überwiegt, kann man sie drei verschiedenen Typen zuordnen (Abb. 6, vgl. ALBERTS et al. 2017, CZIHAK 1981, WOLPERT et al. 2007):
-
Haplonten
Bei Haplonten spielt sich fast der gesamte Entwicklungszyklus in der Haplophase (1n). nur die Zygote (= befruchtete Eizelle) besitzt einen diploiden Chromosomensatz (2n). Diese entsteht durch die Verschmelzung zweier Gameten. Die so entstandene Zygote vollzieht danach eine Reduktionsteilung (Meiose). So entstehen wieder 4 haploide Zellen (Gonen). Zu den Haplonten gehören viele Pilze, viele Algen und einige tierische Einzeller, wie die Flagellaten.
-
Diplonten
Die Diplonten sind quasi das Gegenteil der Haplonten. Bei ihnen vollzieht sich die Lebensphase fast ausschließlich in der Diplophase. Nach der Befruchtung erfolgt, anders als bei den Haplonten, keine Reduktionsteilung (Meiose), sondern die Zygote entwickelt sich zum diploiden Organismus. Einige dieser Zellen bilden dann die haploiden Gameten. Neben einigen Algen und Pilzen gehören alle vielzelligen Tiere (Metazoa) und die Ciliata (Wimpertierchen, Einzeller) zu den Diplonten.
-
Diplo-Haplonten
Die Diplo-Haplonten (auch Haplo-Diplonten genannt) haben eine Zwischenstellung zu den zwei oben genannten Typen. Es sind Lebewesen, bei deren Fortpflanzung abwechselnd haploide und diploide Generationen auftreten. Hierzu gehören die meisten Pflanzen. Kennzeichnend für Diplo-Haplonten ist, dass sowohl in der haploiden wie in der diploiden Phase Mitosen erfolgen. Die haploide Generation pflanzt sich geschlechtlich fort, indem sie weibliche und männliche Gameten bildet. Sie wird deshalb als Gametophyt bezeichnet. Durch die Vereinigung zweier Gameten verschiedenen Geschlechts und Befruchtung entsteht eine diploide Zygote. Die Zygote wächst zum sogenannten Sporophyten. Die Sporophyten bilden nach einer Reduktionsteilung (Meiose) haploide Sporen, die sich ungeschlechtlich Fortpflanzen. Aus diesen halpoiden Sporen wächst der haploide Gametophyt und der Zyklus beginnt von Vorne. In der Evolutionsgeschichte der Pflanzen kommt es dabei zur Reduktion des haploiden Gametophyten. Bei den Moosen ist der Gametophyt die dominante Phase (nämlich die grüne Moospflanze). Schon bei den Farnen ist der diploide Sporophyt dominant und bei den Samenpflanzen ist die eigentliche Pflanze der diploide Sporophyt, während der haploide Gametophyt auf wenige Zellen reduziert ist (männlicher Pollenschlauch, bestehend aus drei Zellen und weiblicher Emybrosack in der Samenanlage, bestehend aus 7-8 Zellen).
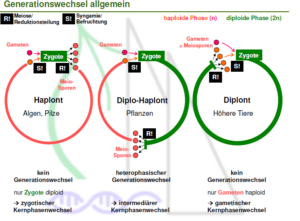
Abb. 6: Kernphasenwechsel. Generationswechsel bei Haplonten, Diplonten und Haplodiplonten. Quelle: Repetico
Typen von Keimzellen (Gameten)
Bei einer Meiose entstehen aus einer diploiden Zelle vier haploide. Doch wenn man sich die Gameten näher anschaut, sehen sie nicht immer gleich aus. Die beiden sich befruchtenden Gameten gehören zwei verschiedenen, als Geschlechter (Sex) bezeichneten Typen an. Entsprechend gibt es auch zwei unterschiedliche Gametentypen. Die kleineren bzw. beweglichen Gameten sind die männlichen Gameten, die größeren bzw. unbeweglichen Gameten bezeichnet man als weiblich. Die Organismen, die in der Lage sind entsprechende Gameten zu bilden werden als männlich oder weiblich eingestuft. Bei der Befruchtung kann stets nur ein Gamet des einen Typen mit dem anderen verschmelzen. Die Gameten sind also die Ursache der bipolaren Zweigeschlechtlichkeit, was sich durch ihre funktionelle Bedeutung wie auch ihre chemischen Struktur, Zelloberfläche etc. ergibt. Darüber hinausgehende morphologische Unterschiede können bei einigen Organismen fehlen. Abb. 7 zeigt die unterschiedlichen Gametentypen.
Sehen die Keimzellen morphologisch (aber nicht zwingend chemisch oder funktionell!) ähnlich, spricht man von einer Isogamie. Im Falle einer solchen Isogamie werden die beiden, nur an ihren funktionellen Eigenschaften unterscheidbaren Sorten willkürlich als + und – bezeichnet, sofern nicht ein Verhaltensunterschied bei der Einleitung der Befruchtung eine Identifizierung des Geschlechtes erlaubt. Lassen sich die Gameten anhand ihres Aussehens unterscheiden, spricht man von Anisogamie. Sind beide Gameten begeißelt unterscheidet man zwischen männlichen Mikrogameten und weiblichen Makorgameten. Ist nur ein Gametentyp begeißelt und damit beweglich, wird dieser – unabhängig von seiner Größe – als männlich definiert. Sind beide Eigenschaften kombiniert, so sind die kleinen und beweglichen Gameten die männlichen Spermatozoen (bei Tieren Spermien genannt) und die großen unbeweglichen Gameten sind die weiblichen Eier. Diese Kombination bezeichnet man auch als Oogamie. Sie kommt bei allen Tieren (also auch uns Menschen) vor. Der Grund weshalb Eizellen übrigens größer sind als Spermien, liegt in einer Sonderfunktion der Meiose bei Eizellen. Hier entstehen zwar auch aus einer diploiden Mutterzelle 4 haploide Zellen. Aber drei dieser haploiden Zellen werden zu sogenannten Polkörpern. Die Teilung der 4 Zellen ist inäqual. Von einer großen Zelle wird nur eine kleine Zelle abgegliedert, die sich dann entweder auflöst oder nochmal teilt. So entsteht eine sehr große Eizelle und 3 sogenannte Polkörper, die funktionslos sind. Die Produktion unterschiedlicher Gameten geht natürlich einher mit der Ausbildung unterschiedlicher Strukturen ihrer Erzeuger: Das gilt sowohl für primäre Geschlechtsorgane, sowie die sekundären. Im Extremfall bildet sich ein regelrechter Sexualdimorphismus aus, bei dem Männchen und Weibchen so unterschiedlich aussehen, als ob sie anderen Arten angehören könnten (vgl. ALBERTS et al. 2017, CZIHAK 1981, WOLPERT et al. 2007). Da in der Natur also auch immer zwei unterschiedliche Gametentypen pro Art ausgebildet werden, ist auch hier die Einteilung in zwei Geschlechter sinnvoll. Ein drittes Geschlecht würde biologisch nur dann Sinn machen, wenn es triploide Arten gibt, die aus drei unterschiedlichen Keimzellen hervorgehen.
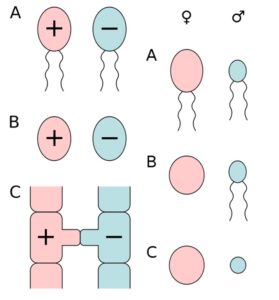
Abb. 7: die unterschiedlichen Gametentypen. Linke Spalte: A) aktiv bewegliche Isogameten, B) unbewegliche Isogamten, C) Gametangiogamie: Befruchtung ohne Gameten bei den Jochpilzen. Rechte Spalte: A) aktiv bewegliche Anisogameten, B) Eizelle und Spermium (Oogamie), C) unbewegliche Anisogameten (Eizelle und Spermatium). Quelle: wikipedia
Zwar gibt es sogenannte polyploide Arten, bzw. Zellen (d. h. ihr Chromosomensatz ist mehr als doppelt vorhanden). Diese sind aber hautsächlich die Ursache einer fehlerhaften Meiose, bei dem der Spindelapparat nicht oder unvollständig ausgebildet wird und so die homologen Chromosomen nicht gleichmäßig verteilt werden können. So können z. B. diploide Gameten entstehen. Doch auch hier haben wir nur zwei Keimzellentypen.
Im dritten Teil befassen wir uns mit der Geschlechtsbestimmung, den genetischen Unterschieden zwischen den Geschlechtern, dem Problem der Intersexualität und Zwittrigkeit und den Argumenten von Fausto-Sterling und Heinz-Jürgen Voß, die von mehr als zwei Geschlechtern ausgehen.
Literatur
ALBERTS, B. et. Al (2017): Molekularbiologie der Zelle. Wiley-VCH; Auflage: 6.
CZIHAK, G., LANGER, H., ZIEGLER, H. (1981): Biologie: Ein Lehrbuch (Springer-Lehrbuch). Springer-Verlag, Auflage 2
DEGROUCHY, J. 1987: Chromosome phylogenies of man, great apes, and Old World monkeys. Genetica. 1987 Aug 31;73(1-2):37-52. https://www.ncbi.nlm.nih.gov/pubmed/3333352
FAN, Y., NEWMAN, T., LINDARDOPOULOU, E. & TRASK, B. J. (2002): Gene Content and Function of the Ancestral Chromosome Fusion Site in Human Chromosome 2q13–2q14.1 and Paralogous Regions. Genome Res. 2002 Nov; 12(11): 1663–1672. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC187549/
LANE, N. 2009: Life Ascending: The Ten Great Inventions of Evolution. Profile Books
MARGULIS, L. & SAGAN, D. 1986: Origins of Sex: Three Billion Years of Genetic Recombination. Yale University Press
WEISMANN, A. 1883: Die Entstehung der Sexualzellen bei Hydromedusen. Zugleich ein Beitrag zur Kenntnis des Baues und der Lebenserscheinungen dieser Gruppe. Verlag Gustav Fischer
WEISMANN, A. 1886: Die Bedeutung der sexuellen Fortpflanzung für die Selektionstheorie. Verlag Gustav Fischer
WOLPERT, L et al. (2007): Principles of Development: Das Original mit Übersetzungshilfen: Das Original Mit Übersetzungshilfen
Video-Beiträge:
The Origin of Sexual Reproduction
