
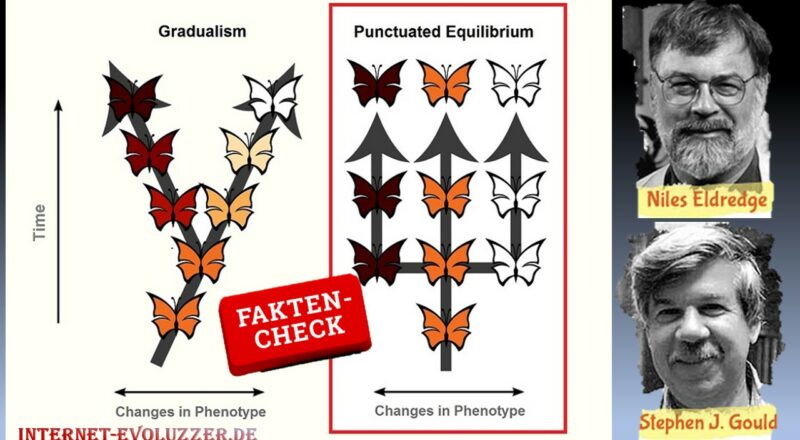
Verläuft Evolution langsam, unmerklich und in kleinen Schritten oder macht die Natur doch große Sprünge und Evolution verläuft schneller als gedacht?
Darwin, wie übrigens die meisten Menschen im viktorianischen England, war Teil einer Kultur, die an Fortschritt und allmähliche Veränderungen glaubte. Von den Revolutionen in Frankreich und Amerika erschüttert, vertraten die Briten stattdessen die Auffassung, dass langsame, stetige Veränderungen das beste Modell für gesellschaftliche Veränderungen und Reformen seien. Als enger Freund und Schüler von Charles Lyell wollte Darwin Lyells Ansatz der allmählichen, uniformen Umwandlung der Erde auf die Biologie ausweiten. Darwin war also ein Anhänger des Gradualismus. Darwins Freund und Förderer Thomas Huxley mahnte jedoch, dass Gradualismus kein notwendiger Bestandteil der Evolutionstheorie sein muss (vgl. Prothero 2017: S. 85-86).
Was sagt der Fossilbericht?
Text als pdf (deutsch)
Im Laufe des Jahrhunderts nach Darwin versuchten Paläontologen zu belegen, was Darwin erwartet hatte. Einige wenige Beispiele für sich scheinbar allmählich entwickelnde fossile Sequenzen wurden dokumentiert, aber im Allgemeinen wurden nicht viele gefunden (Abb. 1).
Hinzu kam: in der Mitte des 20. Jahrhunderts dominierten in der Evolutionstheorie die Gedanken der neu aufstrebenden Genetik und wichtige Felder der Evolutionsbiologie, wie Entwicklungsbiologie und Paläontologie wurden in den Hintergrund gedrängt. Man stellte sich vor, dass evolutionäre Änderungen jederzeit während der Existenz einer Art stattfinden können. Erscheinen in der paläontologischen Überlieferung dennoch Formen mit neuen, scheinbar plötzlich („sprunghaft“) entstandenen Merkmalen, dann erklären sie dies mit Lücken in der fossilen Aufzeichnung.
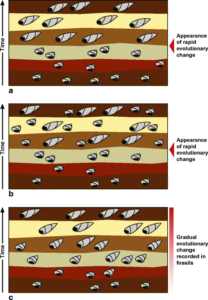
Abb. 1: Hypothetische stratigraphische Aufzeichnungen mit fossilen Schneckenhäusern. (a) Ein punktuelles Muster, bei dem sich eine Art anscheinend schnell zu einer neuen Art entwickelt hat. (b) Ein punktuelles Muster, bei dem eine Abstammungslinie schnell zwei Tochterarten hervorgebracht zu haben scheint, von denen die eine eine mit dem Vorfahren identische Form und die andere eine andere Form aufweist. (c) Ein Muster, das einen langsamen und stetigen evolutionären Wandel aufzeigt.
Eine Gruppe von Paläontologen um Stephen Jay Gould und Niles Eldredge (Abb. 2) legte Anfang der 70er-Jahre des 20. Jahrhunderts eine Alternative vor, mit der Begründung, dass fast nirgendwo graduelle Änderungen zu sehen sind (Eldredge & Gould 1972, Gould & Eldredge 1977).

Abb. 2: Niles Eldredge und Stephen J. Gould
Das Fehlen dieser graduellen Änderungen sei nicht das Ergebnis eines lückenhaften Fossilberichtes, sondern die paläontologischen Daten deuten darauf hin, dass Evolution „punktuell“ verläuft. Demnach hätte eine gerade entstandene Art ihren charakteristischen Phänotyp schnell entwickelt, sich danach aber im Lauf ihrer weiteren Existenz nicht wesentlich verändert.
In den Fällen, in denen die fossile Überlieferung gewisse Muster eines langfristigen Trends erkennen lässt (z. B. die Größenzunahme einer Stammgruppe), lässt sich oft folgendes Muster erkennen: Langfristig existierende Arten, mit denen nicht viel passierte (diese Phase nennen wir „Stasis“) können plötzlich verschwinden und werden durch eine oder mehrere nah verwandte Arten ersetzt (Abb. 3).
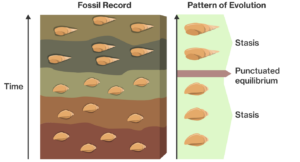
Abb. 3: In den unteren drei Schichten sind Fossilien in Gesteinsschichten zu sehen, die die Einheitlichkeit – die biologische Stasis – über mehrere geologische Zeiträume hinweg zeigen. Irgendwann kam es zu einer raschen Veränderung der Art, und die ursprüngliche Molluske entwickelte sich zu einem anders geformten Organismus mit einer zylindrischeren Schale, wie in den oberen Schichten zu sehen ist. Dies ist ein Beispiel für ein punktuelles Gleichgewicht, d. h. zwei Perioden des Stillstands, die durch rasche evolutionäre Veränderungen unterbrochen werden.
Während ihrer „ereignislosen“ Existenz sind die Arten zwar nicht völlig unveränderlich, aber es sind keine eindeutigen Trends erkennbar – ihre morphologischen Werte oszillieren chaotisch um den Anfangszustand. Es entsteht der Eindruck, dass bedeutende morphologische (und wohl auch ökologische) Veränderungen mit der Aufspaltung der alten Arten in Tochterarten verbunden sind.
Gould und Eldredge nennen ihre Theorie „punctuated equilibrium“, zu Deutsch: „Theorie des unterbrochenen Gleichgewichts“, auch oft als Punktualismus abgekürzt (Abb. 4).
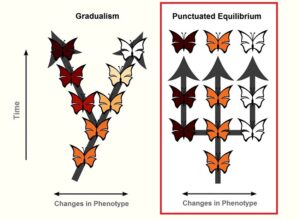
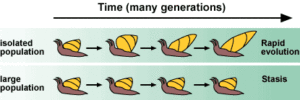
Abb. 4: Unterschied zwischen Punktualismus und Gradualismus
In der Paläontologie gibt es eine Vielzahl solcher Beispiele.
Wie eine solche fossile Überlieferung üblicherweise aussieht, zeigen die evolutionären Beziehungen von 18 Arten mariner Moostiere (Ectoprocta = Bryozoa i.e.S.) der Gattung Metrarabdotus während der letzten 20 Millionen Jahre. Nichts weist hier auf größere morphologische Veränderungen innerhalb der Arten hin, obwohl es manche von ihnen geschafft haben, sich in der fossilen Überlieferung über mehrere Millionen Jahre zu erhalten (Abb. 5).
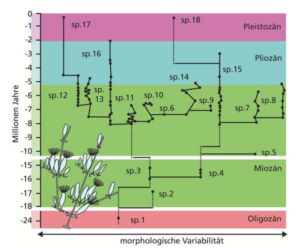
Abb. 5: Evolutionäre Beziehungen der achtzehn Arten (sp. 1 bis sp. 18) mariner Moostierchen (Ectoprocta = „Bryozoa“) der Gattung Metrarabdotus während der letzten 20 Millionen Jahre. Manche Arten haben sich in der fossilen Überlieferung über mehrere Millionen Jahre erhalten, doch es fand keine bedeutende morphologische Differenzierung statt – zu erkennen an den mehr oder weniger vertikal verlaufenden Stammlinien über der Morphologieachse. (Nach Cheetham 1986)
Einige Biologen versuchten, diese Stasis mit Mechanismen wie der stabilisierenden Selektion (Selektion gegen die Extreme einer Population, Verstärkung der mittleren Tendenz) zu erklären, aber dies erklärt nicht, wie einige fossile Populationen über Millionen von Jahren gut dokumentierter klimatischer Veränderungen (sicherlich ein starker Selektionsdruck) unverändert überdauern, wie von Prothero und Heaton (1996) und Prothero (1999a) dokumentiert. Wie Gould (1980, 2002) feststellte, deutet das Fortbestehen fossiler Arten über Millionen von Jahren intensiven Selektionsdrucks darauf hin, dass sie nicht unbegrenzt durch Selektion formbar sind, sondern stattdessen über eine Integrität oder eine Art internen homöostatischen Mechanismus verfügen, der den meisten externen Selektionen widersteht. Dies ist ein radikaler Gedanke für die Evolutionsbiologie, der immer noch heftig umstritten ist.
Für Paläontologen war diese Tatsache jedoch nicht so verblüffend wie für Biologen. Obwohl die Paläontologen ein Jahrhundert lang versucht hatten, den Darwinschen Gradualismus in den Fossilien zu finden, war es die Stasis und das abrupte Auftreten fossiler Arten, die die Biostratigraphie so gut funktionieren ließen (Abb. 6). Wenn sich alles allmählich entwickeln würde, gäbe es ein großes Problem, wie man eine sich allmählich verändernde Abstammungslinie in nicht willkürliche Artensegmente aufteilen könnte. Alle Paläontologen wussten, dass die praktische Welt des Fossiliensammelns selten ein solches Dilemma mit sich brachte. Die älteste Wahrheit der Paläontologie besagt, dass die überwiegende Mehrheit der Arten im Fossilbericht vollständig ausgebildet ist und sich während der langen Zeit ihres späteren Bestehens nicht wesentlich verändert (die durchschnittliche Zeitspanne für die Arten der wirbellosen Meerestiere liegt bei 5 bis 10 Millionen Jahren). Mit anderen Worten: ein geologisch abruptes Auftreten, gefolgt von einer anschließenden Stabilität.
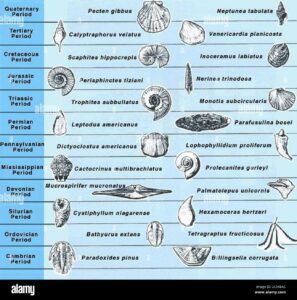
Abb. 6: Biostratigraphie mit Leitfossilien
Der Punktualismus gehört zu den am heißesten debattierten Theorien in der Paläontologie bzw. Evolutionsbiologie – aber auch zu jenen, die am meisten missverstanden werden und besonders von Evolutionsgegnern stark verzerrt oder verfälscht werden. Hier soll es darum gehen die Theorie des unterbrochenen Gleichgewichts in den richtigen Kontext der Evolutionsforschung zu stellen. Denn es handelt sich dabei nicht um eine antievolutionäre Theorie, sondern bezieht sich auf Evolutionsgeschwindigkeiten.
Punktualismus und Saltationismus
Die Theorie wendet sich gegen den so genannten phyletischen Gradualismus, welcher nach Gould und Eldredge eine langsame und mit konstanter Geschwindigkeit fortschreitende Transformation biologischer Arten annimmt. Im Gegensatz dazu wechselt im Punktualismus ein mit „Stasis“ (Stillstand) bezeichneter zeitlicher Abschnitt, in dem Arten nur ein geringes Ausmaß an morphologisch auffälliger Veränderung erkennen lassen, mit schnellem Wandel während ab („schneller“ Wandel ist dabei, da er sich auf geologische Zeiträume bezieht, nicht unbedingt „schnell“ für menschliche Verhältnisse im Sinne des Alltagsgebrauches).
Des Weiteren setzt die Theorie des Punktualismus nicht unbedingt große und plötzliche („sprunghafte“) Änderungen voraus: Damit ist der Punktualismus kein Saltationismus. Ein bekannter Vertreter des Saltationismus war der Entwicklungsbiologe Richard Goldschmidt (Goldschmidt 1940). Den Saltationisten zufolge verlaufen die größeren Evolutionsänderungen von einer Generation zur anderen sprunghaft durch Makromutationen (Abb. 7). Durch solche „Hopeful Monsters“ sollten morphologische Änderungen, z. B. die Entwicklung von Flügeln aus Armen, innerhalb von wenigen Generationen stattfinden. S. J. Gould hat dieser Interpretation widersprochen und betont, dass der Punktualismus im Rahmen der Synthetischen Evolutionstheorie steht und hat einen Bezug des Punktualismus zu Goldschmidts Hypothese verneint. Die Änderungen, die im Augenblick des Umbruchs verlaufen, müssen nicht größer sein als die von den Gradualisten angenommenen Mutationen. Der Unterschied liegt nur in der zeitlichen Verteilung der Evolutionsereignisse, nicht in ihrer „Größe“. Während die Gradualisten relativ konstante Mutationsraten annehmen, gehen die Befürworter des Punktualismus davon aus, dass lange Perioden der Ereignislosigkeit von Phasen mit Evolutionsänderungen durchsetzt sind (vgl. auch Elsberry 1997, Zrzavý et al. 2004: S. 404).

Abb. 7: Eine Drosophila-Mutante (Bithorax-Mutante) mit zwei Thoraxen und vier Flügeln als Beispiel einer Makromutation
Punktualismus und Artbildung
Dass der Fossilbericht eine punktuelle Evolution, statt einer konstanten, aufzeigt, kann Ursache des Artbildungsprozesses sein. In den 1940ern und 50ern entwickelten Biologen wie Ernst Mayr wie neue Arten entstehen können. Darwin war davon ausgegangen, dass man zur Erklärung des Ursprungs der Arten (wie in seinem Titel) nur die Umwandlung von Abstammungslinien benötigte. Mitte des 20. Jahrhunderts war jedoch klar, dass das eigentliche Problem bei der Artbildung die Aufspaltung von Abstammungslinien in zwei oder mehr neue Arten war. Feldbiologen, die das Zusammenspiel von Arten in der Natur beobachteten, stellten fest, dass Arten durch reproduktive Isolation definiert sind, d. h. verschiedene Arten können sich nicht erfolgreich miteinander kreuzen. Ernst Mayr (1942) stellte bei der Untersuchung von Vögeln in Neuguinea fest, dass die meisten Arten unterschiedliche, sich nicht überschneidende geografische Verbreitungsgebiete hatten und dass Inseln oft eigene Arten beherbergten. Daraus, schlug er das Modell der allopatrischen Artbildung vor (Abb. 8). Neue Arten entstehen durch die Isolation einer Population in mehrere Teilpopulationen. Diese können durch die Isolation, z. B. geographische Barrieren wie ein Fluss oder Gebirge, ihre Gene nicht mehr untereinander austauschen; die Populationen sind isoliert, akkumulieren verschiedenen Mutationen und entwickeln sich unabhängig voneinander. Verschwinden Isolationsbarrieren und die Teilpopulationen begegnen sich wieder, pflanzen sich aber nicht untereinander fort, entstehen neue biologische Arten.
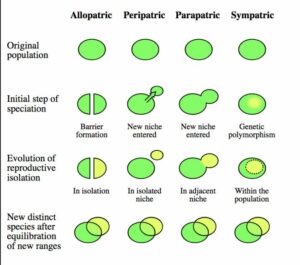
Abb. 8: Formen der Artbildung
In den 1940er Jahren hatten Studien über Züchtung und Genfluss gezeigt, dass große Populationen gegen evolutionäre Veränderungen resistent sind, weil die ungewöhnlichen Gene jeder neuen Variante durch Kreuzung mit den normalen Mitgliedern der übrigen großen Population schnell verdünnt werden. Aber kleine Populationen können sich in relativ kurzen Zeiträumen dramatisch verändern. Wenn zum Beispiel eine Population (oder auch nur ein einziges schwangeres Weibchen) einen isolierten Ort wie eine Insel erreicht, werden alle seltenen genetischen Mutationen bald dominant, da alle Inselbewohner ihre Nachkommen sind. Dieser Effekt ist nicht auf Inseln beschränkt. Kleine Populationen auf dem Festland können sich genetisch unterscheiden, wenn sie sich nicht mit anderen Populationen vermischen. Religiöse Sekten wie die Amish in Pennsylvania weisen beispielsweise eine hohe Häufigkeit ungewöhnlicher Mutationen auf, weil sie eine kleine isolierte Population, die stark durch Inzucht geprägt ist, sind und nur wenige Nicht-Amische konvertieren und in die amische Bevölkerung einheiraten, um den Genpool aufzufüllen. Aus diesen Studien schloss Mayr (1942), dass kleine isolierte Populationen am Rande der Hauptpopulation (periphere Isolate) die wahrscheinlichste Quelle für neue Arten sind.
Mitte der 1950er Jahre wurde dieses Modell der allopatrischen Artbildung von fast allen Biologen weitgehend akzeptiert (mit Änderungen, da immer mehr Ausnahmen auftraten – Ein Überblick zum Thema Artbildung siehe Kubi 2019).
Überraschenderweise kamen aber die Erkenntnisse der allopatrischen Artbildung bei den Paläontologen noch nicht an. Es waren erst Gould und Eldredge, die das Modell der allopatrischen Artbildung auf den Fossilbericht anwendeten (Abb. 9a-g).

9a: 1. Stasis: Eine Molluskenpopulation befindet sich im Stillstand, lebt, stirbt und wird alle paar hunderttausend Jahre versteinert. Nach diesen Fossilien zu urteilen, scheint es kaum eine beobachtbare Evolution zu geben.
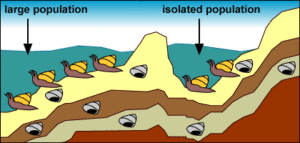
9b: 2. Isolation: Ein Rückgang des Meeresspiegels bildet einen See und isoliert eine kleine Anzahl von Mollusken vom Rest der Population.
9c: 3. Starke Selektion und schneller Wandel: Die kleine, isolierte Population erfährt aufgrund der neuartigen Umwelt und der geringen Populationsgröße eine starke Selektion und rasche Veränderungen: Die Umwelt in dem neu entstandenen See übt einen neuen Selektionsdruck auf die isolierten Mollusken aus. Außerdem bedeutet ihre geringe Populationsgröße, dass die genetische Drift ihre Evolution beeinflusst. Die isolierte Population unterliegt einem schnellen evolutionären Wandel. Dies beruht auf dem Modell der peripatrischen Speziation. Ein Diagramm, das zeigt, dass sich die große Population über viele Generationen hinweg nicht verändert, während die isolierte Population eine verlängerte Schale entwickelt.
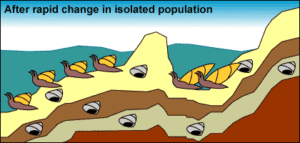
9d: 4. Keine Konservierung: Fossilien, die Übergangsformen repräsentieren, sind aufgrund ihrer relativ geringen Populationsgröße, des schnellen Tempos der Veränderungen und ihrer isolierten Lage nicht erhalten. Schnecken auf dem Meeresboden. Die isolierte Population hat längliche Schalen, die jedoch in den darunter liegenden Schichten nicht erhalten sind.

9e: 5. Wiederansiedlung: Der Meeresspiegel steigt und bringt die isolierten Mollusken mit ihrer Schwesterpopulation wieder zusammen. Bei höherem Meeresspiegel können die beiden Schneckenpopulationen wieder zueinander finden.
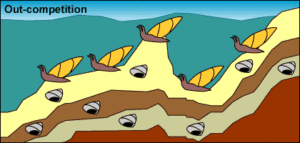
9f: 6. Ausbreitung und Stasis: Die isolierte Population dehnt sich in ihr früheres Verbreitungsgebiet aus. Eine größere Populationsgröße und eine stabile Umwelt machen evolutionäre Veränderungen weniger wahrscheinlich. Der ehemals isolierte Zweig des Weichtierstamms kann seine Vorgängerpopulation verdrängen, so dass diese ausstirbt. Im Unterwasserbereich gibt es nur Schnecken mit länglichen Schalen. Die stratigraphischen Schichten enthalten nur kurze Schalen.

9g: 7. Erhaltung: Eine größere Populationsgröße und ein größeres Verbreitungsgebiet bringen uns zurück zu Schritt 1: Stasis mit gelegentlicher Fossilerhaltung. Im Unterwasserbereich gibt es nur langgestreckte Schneckenhäuser. Die oberste stratigraphische Schicht enthält längliche Schneckenhäuser. Die unteren Schichten enthalten nur kurze Schneckenhäuser.
Wenn das Modell der allopatrischen Artbildung auf den Fossilbericht angewandt wird, sollten wir keine Artbildung in den Fossilien der Hauptpopulation erwarten. Stattdessen sollte die Artbildung in kleinen, peripher isolierten Populationen stattfinden, die kaum eine Möglichkeit haben Fossilien zu werden. Darüber hinaus haben alle Daten aus der Biologie gezeigt, dass dieser Prozess der Artbildung typischerweise in wenigen Hunderten bis Tausenden von Jahren abläuft, was für Paläontologen ein geologischer Wimpernschlag ist. Der Altersunterschied zwischen zwei Schichtebenen beträgt oft viele tausend Jahre. Daher ist nicht zu erwarten, dass die allmählichen Übergänge zwischen den Arten sehr oft erhalten bleiben; stattdessen erwarten wir neue Arten, wenn sie nach ihrer Isolierung und Artbildung wieder in die Hauptpopulation einwandern.
Mit anderen Worten, sie würden plötzlich im Fossilbericht auftauchen. Sobald sie sich etabliert haben, würde die Artbildungstheorie vorhersagen, dass die Hauptpopulation stabil bleibt und sich im Laufe der Zeit nicht allmählich verändert, sondern dass ständig neue Arten in der Peripherie auftauchen und in das Stammgebiet zurückwandern.
Punktualismus und Kreationismus
Natürlich reiben sich Kreationisten kräftig die Hände und missinterpretierten die Theorie des Punktualismus als ein Beweis dafür, dass Evolution nicht stattfindet und die Arten „plötzlich“ erschaffen worden sind. Dabei schrecken sie auch nicht davor zurück, Zitate aus dem Zusammenhang zu reißen und werden so interpretiert, dass Gould und Eldredge behaupten, es gäbe keine Übergangsformen oder dass die Fossilienaufzeichnungen keine Beweise für die Evolution liefern! Typischerweise ziehen diese „Zitatjäger“ einen einzelnen kurzen Abschnitt aus einem längeren Zitat heraus, der genau den gegenteiligen Eindruck von dem vermittelt, was der Autor wirklich gesagt hat. Eine solche Praxis deutet darauf hin, dass die Kreationisten entweder nicht lesen können, das gesamte Zitat nicht verstehen oder absichtlich versuchen, ihre eigenen Leser zu täuschen, indem sie behaupten, Gould und andere hätten etwas gesagt, was in Wirklichkeit das Gegenteil von dem ist, was gemeint war (was bedeutet, dass die Kreationisten unehrlich und betrügerisch sind)!
Die Seite Talk-Origins.org dokumentiert eine Vielzahl solch „Quote-Mining“ durch die Kreationisten, es ist in der Videobeschreibung verlinkt. (http://www.talkorigins.org/faqs/quotes/mine/contents.html)
Aber widerlegt das Konzept des Punktualismus die Existenz von Übergangsformen im Fossilbericht? Nein!
Wichtig ist zu verstehen, was Evolutionsbiologen mit Übergangsformen meinen. Aufgrund der allopatrischen Artbildung und der Seltenheit, dass Fossilien überhaupt entstehen können, sind Art-zu-Art-Übergange vergleichsweise gering. Über 99% aller Lebewesen sind im Verlauf der Erdgeschichte ausgestorben und nur ein winziger Bruchteil davon wird zum Fossil. Um mal mit Zahlen zu bombardieren: heute sind etwa 2 Mio. Tier- und Pflanzenarten beschrieben worden. Die tatsächliche Zahl dürfte aber fast astronomisch größer sein, denn regelmäßig werden neue Arten entdeckt und beschrieben. Wenn wir davon ausgehen, dass 10 Mio. Arten unsere heutige Erde bevölkern und im Verlauf der Erdgeschichte 99% aller Arten ausgestorben sind, brachte die Evolution 1 Milliarde unterschiedlicher Arten hervor! Mindestens! Doch wie viele dieser ausgestorbenen Arten kennen wir als Fossilien? Bis 1999 sind rund 250.000 fossile Arten wissenschaftlich beschrieben worden (Prothero 1999b), runden wir – über 20 Jahre später – auf 300.000 auf. Das heißt nur ein Bruchteil aller Arten wird überhaupt zum Fossil und es wird nicht jedes Fossil entdeckt. Folgerichtig, wenn nicht alle Arten fossilisieren und wir nicht alle Fossilien entdecken, ist es sehr vage zu behaupten Fossil X ist der Vorfahre von der heute lebenden Art Y.
Ein wichtiges Konzept ist hier die Unterscheidung zwischen einem linearen und einem kollateralen Vorfahren (Vgl. Prothero 2017: S. 89-92). Ein linearer Vorfahre ist einer unserer direkten Vorfahren: Vater, Mutter, Großeltern, Urgroßeltern und so weiter. Ein kollateraler Vorfahre hat einen gemeinsamen Vorfahren mit uns, aber wir sind nicht die direkten Nachfahren: Onkel, Tante, Großonkel, Großtanten und so weiter. Bei den Fossilien sprechen wir oft davon, dass ein bestimmtes Fossil nicht direkt von einem anderen Organismus abstammt, sondern anatomische Merkmale aufweist, die darauf hindeuten, dass es mit ziemlicher Sicherheit ein kollateraler Vorfahre ist (oder eine Schwestergruppe oder ein nächster Verwandter).
Wenn ein Ahnenforscher meinen Familienstammbaum untersuchen will und meine Cousine und meinen Großonkel ausfindig machen kann, nicht aber meine Eltern, so ist ja nicht abzustreiten, dass sie mit mir verwandt sind. Und nur, weil man meine Eltern nicht findet, heißt es nicht automatisch, dass ich durch einen göttlichen Akt geschaffen wurde. Genauso verhält es sich mit dem Fossilbericht. Wenn wir Fossilien finden und sie verwandtschaftlich in die Nähe heute lebender Arten rücken, sagen wir damit nicht, dass wir genau den Vorfahren entdeckt haben, der direkt zur untersuchten heute lebenden Art führte. Wir sagen lediglich, dass dieses Fossil eine Reihe von Merkmalen mit heute lebenden Arten hat, die auf eine gemeinsame Abstammung hindeuten. So können wir z. B. sagen, dass Vögel von theropoden Dinosauriern abstammen und Archaeopteryx ist dafür ein Beispiel. Wir sagen aber nicht, die Art Archaeopteryx lithographica ist der lineare Vorfahre aller heute lebenden Vögel.
Was wir also durchaus tun können ist, den evolutionären Übergang von Großgruppen nachzuweisen. So gibt es Übergänge zwischen Fischen und Landwirbeltieren, Amphibien und Reptilien, Reptilien und Vögeln, Reptilien und Säugetieren usw. usf. Eigentlich gibt es zwischen jeden Großgruppen im Tierreich eine Menge von Übergangsformen, die eine Brücke zwischen diesen Großgruppen bauen. In einigen Fällen sind sogar Art- zu Art-Übergänge dokumentiert.
Kathleen Hunt hat auf der Seite talkorigins.org zwischen 1994 und 1997 eine Liste fossiler Übergangsformen bei Wirbeltieren erstellt. Der Titel dieser Liste trägt den Namen „Transitional Vertebrate Fossils FAQ“ (Hunt 1997). Bedenkt, dass diese Seite mittlerweile über 20 Jahre alt ist und sich in diesen zwei Jahrzehnten viel in der Paläontologie getan hat, gerade auch was Neuentdeckungen bei Fossilien angeht. Dennoch ist diese Liste eine schöne Quelle, für diejenigen, die Interesse daran haben, solche Übergangsformen zu sehen.
Schwächen des Punktualismus
Jedoch hat die Theorie des Punktualismus auch ihre Grenzen. Die Stasis in fossilem Material kann nur schwerlich enthüllt werden, weil wir nur die fragmentierte Morphologie sehen. Anders ausgedrückt: wir finden von den Fossilien die Knochen, Schalen und Abdrücke, jedoch sehr selten Weichgewebe, Genome, Färbungen oder gar das Verhalten. So können sich Arten über einen langen Zeitraum vielleicht in morphologischer Sicht kaum oder gar nicht verändert haben. Es kann aber durchaus sein, dass sie sich in Färbungen, Verhalten oder molekulargenetisch stark verändert haben – es ist aber im Fossilbericht nicht überliefert. Ein gutes Beispiel zeigt die Untersuchung sog. lebender Fossilien, also Arten, die sich von ihren Vorläufern kaum verändert haben bzw. sehr viele ursprüngliche Merkmale beibehalten haben. Hier sei ein Beispiel erwähnt: die Brückenechse (Sphenodon punctatus, Abb. 10). Sie ist die einzige Art der Familie der Sphenodontidae und lebt auf Neuseeland (Böhme 2004). Ihrer Erscheinung nach erinnert sie an eine Eidechse, doch beide gehören zu verschiedenen phylogenetischen Gruppen innerhalb der Lepidosaura.

Abb. 10: Brückenechse
Echsen wie Warane, Geckos, Skinke, Chamäleons und Eidechsen gehören zusammen mit den Schlangen in die Gruppe der Sqamata. Ihre Schwestergruppe bilden die Sphenodontia (Abb. 11).

Abb. 11: Kladogramm der Lepidosauria
Ein entscheidendes morphologisches Merkmal, was die Sphenodontia von den Squamata unterschiedet ist die Existenz eines unteren Schläfenbogens im Schädel. Diapsiden, zu denen alle Reptilien und auch die Vögel gehören, verfügen ursprünglich über zwei Schläfenfenster im Schädel. Die Schläfenfenster sind durch Schläfenbögen abgetrennt. Im Verlauf ihrer Evolution haben sich diese aber mehrfach umgebildet. Bei den Squamata hat sich der untere Schläfenbogen reduziert, bei den Sphenodontia nicht (Abb. 12).
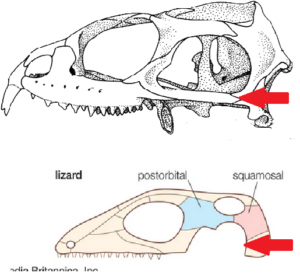
Abb. 12: oben: Schädel der Brückenechse, unten: schematische Darstellung des Schädels einer Echse. Der Pfeil markiert das Vorhanden sein (Brückenechse) oder Fehlen (Echse) des unteren Schläfenbogens.
Die Sphenodontia hatten ihre Blütezeit in der Trias und im Jura – die neuseeländische Brückenechse ist der heute noch einzig lebende Vertreter dieser früher so vielfältigen Gruppe. Morphologisch hat sich die Brückenechse kaum verändert, sie ist vom Skelett her klar als Mitglied der gruppe der Sphenodontia zu erkennen. Ihr Genom weist jedoch eine außerordentlich hohe Geschwindigkeit der molekularen Evolution auf (Hay et al. 2008 – eine relativ neue Studie von Gemmell et al. 2020: S. 408 bemerkt: „We also find that, in contrast to previous suggestions that the evolutionary rate for tuatara is exceptionally fast26, it is the slowest-evolving lepidosaur yet analysed.“). Auch die Schimpansen, die dem gemeinsamen Vorfahren von Schimpansen und Menschen sicherlich stärker ähneln als wir, haben eine schnellere molekulare Evolution als wir Menschen (Zrzavý et al. 2004: S. 406). Das heißt, dass sich molekularbiologisch durchaus bei diesen Arten etwas geändert hat, diese aber keine Auswirkungen auf die Morphologie haben. Genauso kann vermutet werden, dass zwar die fossilen Arten zwar morphologisch in einer Phase der Stasis waren, ihr Genom aber eine starke Evolutionsgeschwindigkeit aufweisen könnte.
Abschluss
Fassen wir zusammen:
Der Punktualismus verbindet die Theorie der allopatrischen Artbildung mit dem Fossilbericht. Die Bildung neuer Arten läuft in kleinen isolierten Populationen schneller ab. Diese haben aber eine sehr geringe Wahrscheinlichkeit zum Fossil zu werden. Das spiegelt sich im Fossilbericht wider.
Der Punktualismus ist kein Saltationismus. Er erfordert keine Makromutationen, bei der neue Merkmale quasi von einer Generation zur nächsten auftreten. Es wird also nicht von einer Generation zu nächsten aus einer Hand ein Flügel. Er sagt lediglich aus, dass sich in kleinen Populationen kleine Mutationen schneller verbreiten können, als in großen.
Der Punktualismus sagt nicht aus, dass im Fossilbericht Übergangsformen von einer Tiergruppe zur anderen fehlen und Evolution unmöglich ist. Im Gegenteil: er bekräftigt, dass durch die Seltenheit zum Fossil zu werden wir Art- zu Art-Übergänge eher selten finden werden. Dies trifft aber auf die Verbindungglieder in der Großgruppensystematik nicht zu.
Literatur
Böhme, W. (2004): Sphenodontida, Schnabelköpfe, Brückenechsen. In: Westheide, W & Rieger, R. (Hrsg.): Wirbel- oder Schädeltiere. Fischer, Stuttgart 2004, ISBN 3-8274-0900-4 (Spezielle Zoologie. Teil 2), S. 354–357
Cheetham A. (1986): Tempo of evolution in a Neogene bryozoan: rates of morphologic change within and across species boundaries. Paleobiology, 12: 199−202.
Eldredge, N., Gould S. J. (1972): Punctuated equilibria: an alternative to phyletic gradualism. Models in Paleobiology, ed. T. J. M. Schopf. San Francisco: Freeman Cooper, pp. 82-115.
Elsberry, W. (1997) „Punctuated Equilibria.“ The Talk.Origins Archive. http://www.talkorigins.org/faqs/punc-eq.html
Gemmell N. J., Rutherford K., Prost S., et al. (2020): The tuatara genome reveals ancient features of amniote evolution. Nature 584(7821):403-409. doi: 10.1038/s41586-020-2561-9.
Goldschmitdt, R. B. (1940). The Material Basis of Evolution, New Haven CT: Yale Univ.Press.
Gould, S. J. (1980): Is a new and general theory of evolution emerging? Paleobiology 6:119-130.
Gould, S. J. (2002): The Structure of Evolutiollary Theon). Cambridge, Mass.: Harvard University Press.
Gould, S. & Eldredge, N. (1977): PE: the tempo and mode of evolution reconsidered. Paleobiology 3:115.
Hay J. M., Subramanian S. , Millar C. D., Mohandesan E., Lambert D. M. (2008): Rapid molecular evolution in a living fossil. Trends Genet. 24(3):106-9. doi: 10.1016/j.tig.2007.12.002.
Hunt, K. (1997): Transitional Vertebrate Fossils FAQ http://www.talkorigins.org/faqs/faq-transitional.html
Kubi, M. (2019): Was ist eine Art? Teil 3/3: Die Entstehung von Arten https://internet-evoluzzer.de/was-ist-eine-art-teil-3-3-die-entstehung-von-arten/
Mayr, E. (1942): Systematics mid the Origin of Species. New York: Columbia University Press.
Prothero, D. R. Heaton, T. H. (1996): Faunal stability during the early Oligocene climatic crash. Palaeogeography, Palaeoclimatology, Palaeoecology 127:239-256.
Prothero, D. R. (1999a): Does climatic change drive mammalian evolution? GSA Today 9(9):1-5.
Prothero, D. (1999b): Fossil Record. In: Ronald Singer (Hrsg.): Encyclopedia of Paleontology. Fitzroy Dearborn Publishers, Chicago, ISBN 1-884964-96-6, S. 491.
Prothero, D. (2017): Evolution – What the Fossils say and why it matters. Second edition. New York: Columbia University Press
Talk Origins: The Quote Mine Project: http://www.talkorigins.org/faqs/quotes/mine/contents.html
Zrzavy,J., , Storch, D., Mihulka, S. (2004): Evolution – Ein Lese-Lehrbuch. Spektrum Akademischer Verlag
